Selection is going to be a key issue for future ground- and space-based observatories. Given lengthy observing times for targets of high interest, we have to know how to cull from our exoplanet catalog those specific worlds that can tell us the most about life in the universe. Recently, Ramses Ramirez (Earth-Life Science Institute, Tokyo Institute of Technology) went to work on the question of habitable zones for complex life, which are narrower than the classic habitable zone defined by the potential for water on the surface. In today’s essay, Alex Tolley looks at Ramirez’ recent paper, which examines the question in relation to the solubility of gases in lipid membranes. What emerges in this work is a constrained habitable zone suited to complex life, with limits Alex explores. The model has interesting ramifications right here in the Solar System, but it also points the way toward constraining the list of planets upon which we’ll apply our emerging tools for atmospheric characterization.
By Alex Tolley

Daggerwrist on Darwin IV. Artist Wayne Douglas Barlowe. Source: Expedition.
Life on Earth, until its last three quarter-billion years, was almost entirely represented by unicellular organisms. As we explored in Detecting Early Life on Exoplanets, biosignatures for microbial life are likely to be far more prevalent than for worlds with complex life. While rocky worlds in the classic habitable zone (HZ) are still relatively few, academic PR departments trumpet every find as “Earth-like”, and a selection of these worlds will be targeted for biosignatures. However, as the number of these worlds increases, scientists will want to distinguish worlds that have a biosphere that can be characterized as more Earth-like, with verdant landscapes and megafauna in the seas and on land.
When the term “Earth-like” is used, the public thinks of a world that looks like Earth, with oceans, continents variously clothed in verdant landscapes, and perhaps most importantly of all, “charismatic megafauna”, the animals that you went to see at the zoo, or watched on David Attenborough’s excellent nature programs. A blue sea lapping on a muddy beach, despite teeming with microbes and other unicellular life, looks dead to the unpracticed eye, which means most of the human population. It is those human-scale animals like the daggerwrist pictured above from Barlowe’s “Expedition: Being an Account in Words and Artwork of the 2358 A.D. Voyage to Darwin IV” that excites the public.
If life is rare, then the classic HZ will have the least constraints, although most of those worlds will still have biospheres populated only with microbes, and fewer probably with unicellular plants and animals. If life is not rare, then there will be a desire to discover true Earth-like worlds with complex life, which may mean limiting the range of the HZ that will allow for such life to flourish.
The classic HZ range is defined by the possibility of liquid water remaining continuously on the surface, warmed by the star’s radiation and an atmosphere of sufficient pressure and with some greenhouse gases. This is because all Earth’s life requires liquid water and this has led to the mantra “Follow the water” for missions in the search for life. Inside the inner HZ limit, there will be a runaway greenhouse that eventually desiccates the planet, like Venus. Towards the outer edge, the atmosphere needs to be increasingly composed of greenhouse gases, particularly carbon dioxide (CO2) until a limit is reached.
For the solar system, the classic HZ lies at about 0.95 AU, inside Earth’s orbit, but excludes Venus, and extends to about 1.67 AU, outside of Mars’ orbit. It is this that offers the possibility of a second genesis and possibility of finding extant life in refuges and in the lithosphere beneath the now inhospitable Martian surface.
Complex, or multicellular, life on Earth emerged less than 1 billion years ago as photosynthesis reduced the CO2 in the atmosphere and replaced it with oxygen (O2). Except for a few recently discovered species, all multicellular life is aerobic and requires a rich O2 atmosphere. It is the much greater energy released by aerobic respiration compared to anaerobic respiration that allows for the energetic lifestyles of multicellular animal life (metazoa). At least for our planet, we believe that the conditions for complex life to survive are constrained; Earth has its own habitable zone limits that are narrower than the classic HZ. The question is, “What might those HZ limits be for complex life, and how does that translate for exoplanets around different stellar types?”
CO2 is one of the main greenhouse gases that extend the outer boundary of the HZ. Nitrogen (N2) also helps extend the outer edge of the HZ although it is not a greenhouse gas but a main constituent of the atmosphere. Are there limits to the pressures of these gases due to effects on complex life that limit the range of the possible HZ for multicellular life living on the planet’s surface?
A new paper by Dr. Ramses Ramirez attempts to answer that question by applying the relationship between the solubility of gases in lipid membranes and their anesthetic potency (see figure 1 below). This theory, a partial explanation for the still imperfectly understood mechanism of anesthesia, is that the solubility of gases in lipid membranes is correlated with their anesthetic potency. Anesthetists must monitor the use of these gases to maintain unconsciousness. Too little and the patient remains conscious of the pain during surgery, too much anesthetic, and the patient stops breathing and dies.
The anesthetic gases are to the bottom right of the chart in figure 1. Nitrous oxide (N2O) is less potent and still used in dentistry (as well as at “nitrous parties”). Less well known is that CO2 also acts as such a gas with solubility similar to N2O. Although physiologically CO2 initially increases breathing rate to flush it out of the lungs, at higher concentrations it then invokes respiratory, and later metabolic, acidosis, which sets in as CO2 dissolved in the blood serum eventually causes cessation of respiration and death. As can be seen in figure 1 below, N2 has low solubility in lipid membranes, 2 – 3 orders of magnitude lower than CO2, and concomitantly similar orders of magnitude lower anesthetic potency.
However, we are probably also familiar with the effects of high-pressure N2 as nitrogen narcosis that is experienced by divers breathing compressed air at depth. The argument is that both CO2 and N2 dissolving in the lipid membranes of cells will cause death if those gas concentrations reach the anesthetic level for complex life.
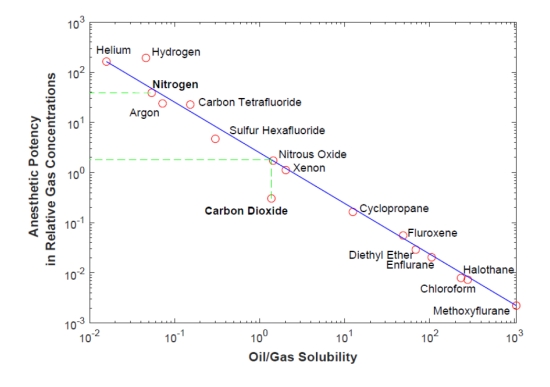
Figure 1: The Meyer-Overton correlation of oil/gas solubility versus anesthetic potential of inhaled gases. Figure recreated from published data. Source Ramirez [1].
Figure 1 above shows the relationship between gases and their anesthetic potential. CO2 solubility is similar to nitrous oxide, while N2 is far less potent and therefore apparently less of a constraint. Note that helium is at the upper end of the range and has low solubility and low anesthetic potency. This is why helium is used to replace N2 when deep diving in soft suits.
While the Meyer-Overton correlation is primarily for humans, it has been shown to apply across several different phyla as it is a physical, rather than physiologic effect. Determining the tolerance limits for CO2 and N2 provides a constraint that limits the HZ to a “Complex Life Habitable Zone (CLHZ).” Dr. Ramirez supports the general applicability of the lipid gas solubility to metazoa from prior experimental work, primarily on mammals, but also with other animals, to suggest that 0.1 bar (1/10th of surface atmospheric pressure or 1.4 psi) of CO2 might be a reasonable, conservative limit for complex life to tolerate CO2. N2 limits are primarily set by experiments for human divers. 2 bar of N2 seems to be the safe limit at which divers do not get narcosis. This is just 10 meters below the surface, a depth even beginner scuba divers can safely operate for short durations. Using upper limits for 0.1 bar CO2 and 2 bar N2, Dr. Ramirez finds that his radiative-convective model (RC) gives an estimated HZ for complex life (CLHZ) of 0.95 – 1.21 AU. Using an advanced energy balance model (EBM) that allows for different temperatures on the Earth’s surface, thus allowing for liquid water at the equator, but not at the poles, this CLHZ is extended from 0.95 – 1.31 AU.
The new outer range for this 2 bar N2 and 0.1 bar CO2 is 1.36 AU using the Energy Balance Model (EBM). This range is shown in figure 2 below not just for Earth, but for a range of main sequence star types. The relative decrease in the CLHZ compared to the HZ is greatest for cooler stars, the type we have most exoplanet examples in the HZ currently.
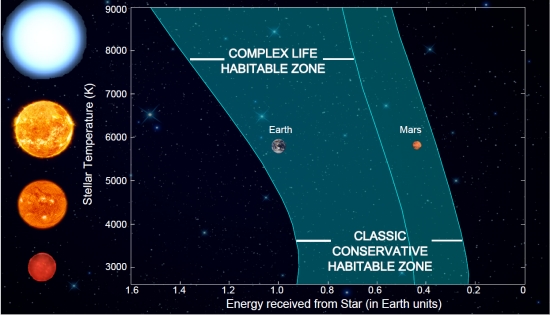
Figure 2. The Complex Life Habitable Zone (CLHZ) for A – M stars (2,600 – 9,000 K) compared to other definitions.The CLHZ is for a 0.1 bar, 2 bar N2 atmosphere which is compared to the classic HZ. While the inner edge of the HZ and CLHZ are the same at 0.95 AU, the outer edge of the CLHZ is now well inside the orbit of Mars. Image source: Ramirez.
Dr. Ramirez compares his results to a similar paper by Dr, Edward Schwieterman that looks at the same problem but through the lens of CO2 chemistry, with the note that carbon monoxide (CO), while not limiting the CLHZ, is toxic and could be limiting to the evolution of complex life [2]. (The CO is created by photolysis of CO2.) Schwieterman uses a 1D radiative-convective climate model for his calculations across a range of CO2 levels. Schwieterman does not investigate higher N2 pressures which results in his modeling having a narrower CLHZ than Dr. Ramirez’s most comparable modeling. However, the CO toxicity does not appear significant except for planets orbiting cool stars such as M dwarfs.
While both authors attempt to redefine the likely boundaries for the HZ of complex life based on Earth’s biological evolution, only Dr. Ramirez employs the possibility of increasing the N2 pressure to increase the outer limit.
To quote from the paper:
“The CLHZ is slightly wider at the higher N2 pressure because of increased N2-N2 collision induced absorption and a decrease in the outgoing infrared flux, which more than offset an increase in planetary albedo.”
Dr. Ramirez also states:
“I consider how our solar system’s HZ changes if we assume (for the moment) that complex life could evolve to breathe in a hypothetical 5-bar N2 atmosphere. For this sensitivity study, the RC model predicts that such worlds in our solar system can remain habitable at 1.24AU (SEFF = 0.65) whereas atmospheric collapse can be avoided as far as 1.36 AU (SEFF = 0.54) in the EBM (nearly 60% classical HZ width). I find that the additional N2 opacity is sufficient to counter the ice-albedo feedback, allowing for effective planetary heat transfer even at relatively far distances.”
Dr. Ramirez’s 0.1 bar constraint for CO2 should be put in context for life on Earth. CO2 is currently at about 0.04% (0.0056 psi) of the Earth’s atmosphere. Even during the Cambrian period when multicellular animals were rapidly diversifying into phyla, the atmospheric component of CO2 was never more than 1% and it fell fairly continuously during this period. The Great Permian Extinction which saw 90-95% of all complex life become extinct primarily by anoxia in the oceans, the CO2 levels were little more than 0.1% at their peak. [See “Climate Change and Mass Extinctions: Implications for Exoplanet Life”] and figure 3 below. For highly cognitive humans, NASA conservatively stipulated that the highest emergency level of CO2 in the Apollo Command and Lunar modules should be no more than 0.29 psi (0.02 bar) in an atmosphere of 5 psi O2 before cognitive skills become impaired [40]. The Centers for Disease Control and Prevention (CDC) guidelines for CO2 is that 0.04 bar CO2 is immediately dangerous [i].
It should also be noted that the analysis is limited to surface living, air-breathing animals. Bathypelagic organisms, such as oceanic fish may be adapted to tolerate far higher N2 pressures.
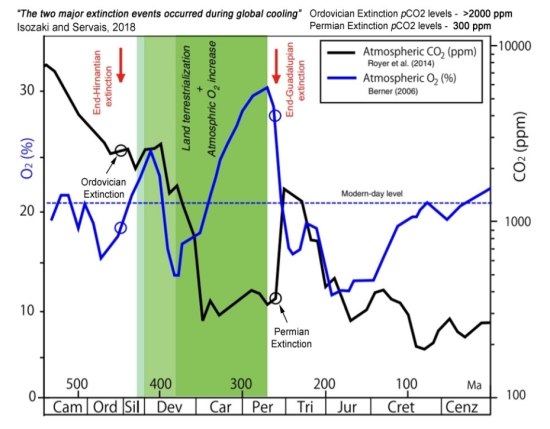
Figure 3. O2 and CO2 levels in the Phanerozoic. [3] While the Permian extinction is associated with a rise in CO2 levels to about 0.1%, and a decline in O2 levels from the Carboniferous, the CO2 levels were far higher at 1% at the start of the Cambrian and still high in the Devonian (the age of fishes).
But what about multicellular organisms other than animals? While Dr. Ramirez acknowledges that complex life includes plants and fungi, not just metazoa (animals), he is unable to address the possible range of CO2 and N2 pressures these complex life forms might be adapted to because there is next to no data on the effect high pressures and concentrations these gases have on plants or fungi, beyond incremental increases in CO2 to experiment on plant photosynthesis limits and productivity. Where we do have data is Earth’s history of complex life that indicates that relatively low levels of CO2 in the atmosphere due to volcanic emissions, and reduced plant life to draw down CO2 and replenish the O2 due to sulfur acid rains and ash-darkened skies, are sufficient to force most species, including plants, to extinction. We do not know what factor or combination of factors is important, nor whether it is primary factors such as anoxia, or n-th order factors that resulted in their final extinction.
Now that the inventory of exoplanets is rapidly increasing, it is certainly time that we start thinking more critically about what sort of life we are looking for and what that might mean for the range of the habitable zone that supports these different life forms. Rather than allowing the widest possible HZ that allows any atmospheric composition and pressure allowing liquid water, we could also be looking for possible constraints that appear required for the sort of surface, air-breathing complex life that will give rise to the charismatic fauna that we have on Earth. Dr. Ramirez has posited one interesting idea for terrestrial complex life that is based on respiration across a range of metazoans which then constrains the atmospheric gas composition and hence the HZ.
As Ramirez’ CLHZ has an outer limit well inside the orbit of Mars, this invites speculation that if Mars ever had any life during its earlier, wetter, period, it did not have complex life. If this model proves correct, while we may find subterranean microbial life on Mars, we will not find metazoan fossils, such as mollusk shells or vertebrate skeletons.
It should be borne in mind that life as a whole maintains Earth’s low CO2 levels to keep the surface temperature equitable for itself, maximizing biodiversity and biomass. While hotter (e.g. the Eocene maximum) and cooler (ice ages) periods upset that equitable temperature, life in concert with much slower geological processes act as a thermostat. It is also the case that biomass and diversity are greatest in the tropical forests and the lowest at the poles. It must have been relatively sparse during the “snowball Earth” period but recovered once the global ice sheets melted. Life has evolved on the Earth as it is, and has biochemistry that matches that requirement.
Today, that requirement is for an atmosphere that has a low CO2 level. On exoplanets, where much higher CO2 levels are needed to keep the planet warm, different biochemistries might develop, and this is a caveat that Ramirez considers for his analysis. However, without examples of such life, we are forced to use Earth’s life as our only model. In a half-billion or so years in the future, as the sun increases its luminosity, the required CO2 level to keep Earth cool enough will be below that needed by plants. A technological species might utilize technology like orbital sunshades or perhaps genetic engineering to maintain life on Earth.
The more important point is that we may be able to provide more granular characterizations of exoplanets. Rather than the binary in or out of the classic HZ for exoplanets and therefore potentially living or not, we can add granularity, such as inside the CLHZ and therefore capable of hosting complex life too. This conclusion does depend on exo-life following our terrestrial biology. If it doesn’t then we have to fall back to the more generous HZ calculations alone.
References
1. Ramirez, Ramses M. “A Complex Life Habitable Zone Based On Lipid Solubility Theory.” Scientific Reports, vol. 10, no. 1, 2020, doi:10.1038/s41598-020-64436-z.
2. Schwieterman, Edward W., et al. “A Limited Habitable Zone for Complex Life.” The Astrophysical Journal, vol. 878, no. 1, 2019, p. 19., doi:10.3847/1538-4357/ab1d52.
3. CO2 and O2 levels in the phanerozoic. Web accessed May 11, 2020. https://notrickszone.com/2018/05/28/2-new-papers-permian-mass-extinction-coincided-with-global-cooling-falling-sea-levels-and-low-co2/
4. Michel, E. L., et al, SP-368 Biomedical Results of Apollo – Chap. 5 Environmental Factors. Accessed from web, May 11th, 2020. https://history.nasa.gov/SP-368/s2ch5.htm



It’s worth bearing in mind that the Schwieterman et al. results about carbon monoxide-based limits to the habitable zone have been challenged as being a bit too vertebrate-centric. As noted by James E. Howell in a comment in RNAAS, hemoglobin is not the only oxygen-transporting protein used by life on Earth:
(emphasis mine)
Cephalopods are among the organisms that use hemocyanin, which is perhaps encouraging for the possibility of intelligent life emerging on worlds with higher carbon monoxide levels.
Andy, thank you for the info on CO and different blood chemistries. I was not aware of this fact and may use it in a follow-up post which is very much in the vein of trying to evaluate atmospheres for more than air-breathing land organisms, which in turn is weighted for vertebrates (which is where most of the data is).
Anyone with good pointers for other organisms, would be welcome.
“Anyone with good pointers for other organisms, would be welcome.”
This might serve as an appetizer.
But of course one with the proclivitymay wander further afield.
Alex and Andy, very interesting and what may be the most common form of intelligent life, Cephalopods. The octopus and cuttlefish both have a large brain, high intelligence and something over look, bioluminescence and optical signaling. We may need to look at what a SETI signal from such a intelligent species may entail, without using electricity, bioluminescence and optical signaling developed to the degree of interstellar communication. They have all these features that an advanced form would need to generate such a signal plus the development of optics with water and water masers. Like the dinosaur raptor that could of become similar to humans if the comet impact had not wiped them out. We need to look at the octopus and and cuttlefish as a good example of what we are up against in trying to communicate with other civilizations. Since many of these cephalopods have had a shell, hard radiation protection is built in. “Binds oxygen via copper, and with distinctive chemistry”, sounds like whole worlds out there are just different from us!
Cephalopods, particularly octopuses (octopoda), have a life span that ends with or before the hatching of the next generation: this excludes passing on of knowledge acquired in one’s lifetime to one’s descendants. Their complex behaviors are selected for by evolution.
The complexity and unique configuration of the cetacean brain and their intelligence are attributed to their unique societal and environmental demands.
Yes the deep dwelling octopus can live up to 5 years if not mating.
The octopus can also rewrite its DNA, which could transfer information learned into the new brood.
The octopus has the weirdest DNA map scientists have ever seen.
“People have been comparing the octopus to an alien life form for years—most notably mollusk expert Martin Wells, grandson of H.G. Wells, who ate his subjects after he experimented on them.”
https://www.dailydot.com/parsec/octopus-dna-map-theyre-aliens-basically/
Researchers have discovered that octopus genomes contain alien DNA.
https://www.snopes.com/fact-check/octopus-dna-origins/
The octopus genome and the evolution of cephalopod neural and morphological novelties.
https://www.nature.com/articles/nature14668
Are octopuses alien? New theory argues Earth was ‘seeded’ by interstellar genetic code.
https://www.foxnews.com/science/are-octopuses-alien-new-theory-argues-earth-was-seeded-by-interstellar-genetic-code
Octopuses or Cephalopods have existed for at least 500 million years. If that count as alien, it must be a record long time to get a proper citizenship on Earth.
I have often joke that Dictyosteliida and Myxogastria must be aliens, the good doctor in the researched used a similar phrase which is not supposed to be taken seriously – but conspiracy peddlers like fox and blogs with a similar intent will of course distort such into something completely different to spin doctor their world view.
Cephalopods have a completely normal genome for a mollusc, it’s only that it have been reshuffled in an unusual way – so lets move on, no aliens to find here!
Also, so far, Paul Davis and his group have failed to find any evidence of a shadow biosphere either. While absence of evidence isn’t proof of evidence of absence, just as with SETI, the odds of presence must be lengthening.
This may be important at an early stage, but if it can be overcome, then technology will help. We use writing, printing and other information storage techniques that allows information to be transmitted without direct communication. If some smart cephalopod could invent some way to store information, then perhaps it could bootstrap the process towards a civilization.
Having said that, I think we make too much of cephalopod intelligence. Yes, they are way smarter than the rest of their phylum, but that is a very low bar. Arthur C Clarke elevated their intelligence stature in his short story: The Shining Ones, about giant squid. Stephen Baxter also had intelligent squid as “astronauts”. Squid and Octopi have had a very long time to evolve, yet they did not progress to the intelligence of some vertebrates.
A basic problem they’ve faced is that water just doesn’t carry that much oxygen, so it’s difficult for fully aquatic lifeforms to have highly energetic metabolisms. The amount of water you’d have to process per minute to capture as much oxygen as a human requires is pretty horrific.
Oh, I doubt that very much, coming from a bag of hot air! How much water does it take to cool a nuclear reactor?
AFAIK, metabolic rates as a function of body size scale across all vertebrates. Fish cellular metabolism is the same as their air-breathing evolutionary descendants which makes me doubt there is any fundamental difference across aquatic and land vertebrates.
Metabolic rate is primarily related to body mass. Fish must move in a very viscous medium, so they tend toswim slowly unless avoiding predators. Think how hard it is to wade through water and how exhausting this is to get a sense of the energy required.
Small insects have a similar problem to fish in that air seems highly viscous to them. That helps in staying airborne for flyers but would restrict their speed. Insects must have low body mass to maintain the metabolic rates to fly in such viscous air.
Ah, I see the earth centered speciesism entering your thoughts!
“WAR of the Worlds” I believe the very first book on alien contact!!!!
You being a biologist, should understand just how different the Cephalopods are from us, but have the ability to outsmart us.
And they have been around for a lot longer then the intelligence of vertebrates.
Once again Sir Alex, you expose me to thoughts that have been batting about my mind. Those thoughts often have a hard time muddling to the surface for me. Thank you for this post, we all get smarter when we listen to each other.
I realise that Octopus parents die before their offspring hatch, but if their offspring were cared for by their fathers, other adults, etc, learning COULD be passed on. Wolves tend each others’ offspring. WE have teachers.
I realise that our cephalopods in fact DO NOT do this. But the point isn’t really about whether our guys do this, it’s about whether ET creatures with physiologies and anatomies quite different to humans, can develop intelligence and potentially culture.
Hi andy..
Argh… I have another comment below this exchange that addresses Howell’s CO comment but was meant to go here… Check it out below.
Since the Galactic Arm passages fit so well to the Geologic Time Scale; associating Oxygenation to anything other than PP lowering CO2 (warming) thus global cooling (Karoo ICE) maybe problematic. Ocean thermal regulation goes further than water heat capacity, as witnessed by the Oligocene global Temperature Notch cunundrem; closely early tracks ICE grounding thermal conductiviy switch after the first ?400k? years.
Your analysis is completely understandable.
A very interesting article. Is the CLHZ going to help us much to limit the number of worlds we think might harbour complex life? Using this new measurement Mars has gone from just inside the HZ to slightly further outside the CLHZ. Any planet near either zone in a different star system will obviously still be of great interest in the future I would think. Still, it gives us a few parameters of importance to work with (N2 and CO2 pressure primarily, and secondarily CO pressure in the atmosphere in question).
Currently we have a binary Hz/non-HZ state -> p(Life)/no Life. As the paper suggests, if we have an abundance of targets, then we might want to focus resources on planets with a p(Complex Life). There will be living planets of microbes that will be harder to gain unambiguous biosignatures. Unicellular photosynthetic life might be easier. But complex life, with terrestrial plants and animals, needed a rich O2 atmosphere should not only be easier but will be far more interesting to the public which in turn will support funding.
Thank you Alex, that helps explain the reasoning. It would be interesting to test the hypothesis Dr. Ramirez. I hope we’re in a position to do so in my lifetime by exploring Mars properly. That would give us an n of 2. From there we would need to get to other star systems to expand the sample size :). Of course if we found complex life forms (arthropod-like or otherwise) in the oceans of one or more of the outer system planets’ moons it would be a huge breakthrough.
Yeah, this comment is brought up a lot. To be fair to Schwieterman et al., I am not sure that Professor Howell’s comment really impacts the Schwieterman et al. analysis much. Mainly, CO actually has little to do with the calculation of HZ limits. This is for two reasons: 1) CO is a weak greenhouse gas and 2) CO atmospheric concentrations are so low that it can be safely ignored in the calculation of HZ limits (and in fact, it is). It is this same reason why other trace gases in Earth’s atmosphere, like N2O and CH4, are ignored in calculations of the “classical” N2-CO2-H2O HZ (and, in turn, the complex life HZ). High concentrations of other gases, like CH4 and H2, are not amenable to complex life as-we-know-it anyway (assuming it requires O2). So, the big greenhouse gases for this application are CO2 and H2O, with the main background gases being N2 (and secondarily, O2).
Ed’s point is that CO *may* be able to build up in these O2-rich atmospheres to levels that are toxic for complex life for HZ planets. However, this potential build up can only occur at all in planets orbiting late (the coolest) M-dwarf stars.
So, it is worth recognizing that CO does not build up to levels that are toxic for complex life in HZ planets orbiting the vast majority of star types (only the coolest M-dwarf stars).
.
Even so, as I mention in my paper, there are bigger issues with the habitability of planets orbiting these coolest M-dwarfs that must be addressed before atmospheric CO can be considered at all. To even have a CO problem to begin with, the planet must be in principle capable of potentially producing complex life (e.g., ocean, some land, high O2, some CO2, N2..etc). However, HZ planets orbiting late M-dwarfs are precisely the ones that have the most issues meeting this condition. The a) superluminosity of pre-main-sequence stars and resultant planetary desiccation, b) high stellar radiation, and c) the prediction that some of these worlds may be ocean worlds (not very “Earth-like”), would all preclude the conditions that could produce planets otherwise capable of complex life, but with a CO build up problem.
Nice summary Alex! I also really like the Daggerwrist figure!
The point of my calculations was to constrain a tighter HZ and eliminate some of the uncertainty with the Schwieterman et al. analysis (which had a big spread of answers for the outer edge) , while also acknowledging that these limits are still ultimately based on our limited understanding of Earth-like life. A tighter HZ is important for astronomical observations that would require more accurate limits in targeting planets that may host complex life.
Just a couple of additional points. The predicted 0.1 bar CO2 and 2 bar N2 respiratory limits are based on newborns, which can be seen as the lowest common denominator. As discussed in my paper, adult animals can withstand higher respiratory pressures of ~0.15 bar CO2 and 3 bar N2, respectively. These numbers are based off of lipid-solubility theory (N2 is ~1/20 as toxic as CO2) and laboratory experiments on all sorts of land animals (emphasizing mammals but also birds, reptiles, insects…etc.), including humans.
One thing in Alex’s nice article that I wish to call out:
“It should also be noted that the analysis is limited to surface living, air-breathing animals. Bathypelagic organisms, such as oceanic fish may be adapted to tolerate far higher N2 pressures.”
It is true that a complex life HZ is limited to air-breathing animals because the HZ is defined with respect to the existence of surface water. Nevertheless, although the complex life HZ can only really assess surface conditions and respiratory limits for air-breathers (and we mostly concerned about atmospheric biosignatures), can we say anything at all about underwater complex life (non-air breathers)? …
So what happens in the ocean? The N2 partial pressure appears to be approximately constant (~0.8 bar) with depth. This occurs because the N2 molar concentration decreases the deeper you go, balancing the concomitant increase in the background H2O pressure. Now, I am no biologist, nor expert in oceans/underwater life, but bathypelagic organisms (e.g. deep sea fishes) are still breathing in seawater with N2 pressures no higher than ~0.8 bar, from my understanding, which would seem to suggest that the aforementioned N2 respiratory limits of 2 – 3 bar might apply here as well.
That said, Alex had sent me a nice article by Scholander and Van Dam (1953) showing that the internal N2 pressure within some of these deep sea fishes can increase beyond that of the ambient seawater by at least a factor of a few. This is because of diffusion exchange that occurs with their special swimbladder. However, what would need to be done is test whether this breathing mechanism would still work if the N2 pressure in the surrounding seawater was elevated above 2 or 3 bar. The question here revolves on answering what the ambient (i.e. environmental) pressure limit is, not the internal pressure limit of an organism. Also, how long would such fish be able to survive under such conditions? Narcosis can be withstood, at least for a limited amount of time, which was the point of the 5-bar N2 sensitivity study in my paper. A swimbladder is evolutionary analagous to the lungs of air-breathers, but it is not quite the same, so testing respiratory limits of underwater organisms is an interesting venue for future work.
How much can lipid membranes be (xeno?)biologically modified to alter gas solubility and permeability? How much can biological systems originally evolve or later adapt to various atmospheric pressures and gas partial pressures?
Efficient oxygen (or equivalent) transport from the exterior of a organism to its interior depths is essential for metabolic systems based on oxidation. Diffusion of gas, even enhanced by venlhlatory mechanisms, imposes a distance constraint that translates into a size constraint for invertebrates in our world. Beyond those constraints, there is transport of oxygen-bearing molecules over substantial distances in addition to mechanisms for the loading and the unloading of the oxygen. Even “the right amount” of water to permit both oceans and dry land is required if air-breathing is to achieve prominence.
Our conceptualization is sadly limited by the sample size of one world.
Good question. This is why the paper and my post point out that this is (or indeed any suitable different evolutionary path) will undermine this Earth-life approach.
There is also the issue of how relevant this is to plant and fungal complex life. I cannot find any data to determine this. This may well be something for astrobiologists to explore.
Thanks Alex and Ramses. An interesting take on the Hab zone paradigm.
I may well have missed something here as I have not had access to the full paper. Apologies if so. As things stand however I am confused by the central premise.
Meyer and Overton independently described their findings around the begining of the last century and these where conflated to become the hypothesis described above . Although seemly logical and robust for six decades it eventually succumbed to scientific falsification by the 1960s and is no longer recognised in the medical world. It was certainly what I was taught at medical school and as a junior doctor whilst working in anaesthetics. Or today having discussed it with an anaesthetic colleague this morning.
The reason for this was based on two findings.
Firstly, that anaesthetic agents bind to proteins as well as and independent of lipids ( in cell membranes for instance ) .
Secondly, that for lipid binding agents there is a cut off point beyond which further binding simply leads to saturation of said binding site . Yet even in such agents the anaesthetic effect continues to increase . All indicating that lipid binding , cell membrane based or not, is not the mechanism of anaesthesia . Current thinking is that cell membrane ion channels and/or intracellular enzymes may be the vehicle for anaesthetic action. But this is still purely hypothetical as the process obviously much more complicated than originally envisaged . As ever ! No one theory is currently held up to anything like the level of orthodoxy of Meyer-Overton at its peak.
The Meyer-Overton effect was also concerned with respect to neuronal cell membranes – particularly in the brain, for the logical reason that any anaesthetic effect would be need to be mediated through the central nervous system and its control on enviromental awareness (from simple touch all the way through to consciousness and sentience ) . Evolution on other planets might possibly lead to some similar chemistry to the Earth. Might. It might even lead to functions analoguous to Earth based organisms – such as photosynthesis and respiration , given the availability and utility of both CO2 and O2 for life . Especially for the terrestrial style HBZ planets considered here.
However the biochemistry, physiology and anatomy of any extra terrestrial organisms, from simple through to complex ( with multicellularity the “simplest” description of the latter ) would be entirely different. So there would be no neurones to mediate enviromental awareness and an entirely different biochemistry to any life. Which would not respond to the concept of anaesthesia as informed and described on Earth. Be it Meyer-Overton or any other ire modern concept.
Hi Ashley. Yeah, I discuss the applicability of the Meyer-Overton correlation (in greater detail in the Supp Info section). It is definitely not perfect for the reasons you discuss. For instance, it could be that anesthetic action occurs in protein binding sites rather than in lipid bilayers (although the latter may be a necessary-but-insufficient condition). Also, the correlation fails for certain complex molecules (e.g. perflourinated compounds).
However, as I showed in my paper, further bolstered from the many experimental studies on animals I cite, the correlation does seem to work reasonably well with the simple gases of interest here, CO2 and N2. This consistency with many standard gases was probably why it had been widely accepted for so long (over a century). Nevertheless, I cite many animal studies, both more recent and older ones, that all still compare their results against the Meyer-Overton correlation. So, it is clearly still used in research circles. As you say though, it is not perfect, but it is still not so bad for this particular application, at least for Earth air-breathing animals exposed to CO2 and N2.
What I assumed here was that alien complex life MAY be similar to Earth’s, exhibiting similar respiratory limits. This could be true because I’ve assume a carbon-based biochemistry that needs water (plus assuming it probably uses something akin to DNA), just like Earth life does. If that happens to be the case for extraterrestrials too, the limits I calculate could generally hold elsewhere (perhaps the many examples of convergent evolution on Earth is the best argument for that).
That is the baseline scenario.
However, as you mentioned, it could be that complex life elsewhere finds different evolutionary solutions for similar problems. In that case, my paper includes some discussion and sensitivity studies addressing this point.
Thanks Ramses.
This article understandably focuses on N2 and CO2 with respect to the fresh “narcosis ” perspective. However i wondered about the parallel impact of methane and especially water vapour on the HBZ, in terms of greenhouse effect for both and in the latter’s case also it’s indirect effect on CO2 partial pressures via the carbon/silicate cycle ?
Best wishes
Ashley
Hi Ashley,
H2O vapor is always included in these HZ definitions, as it is a strong function of temperature. H2O vapor has the biggest impact near the inner edge.
In 2018, we wrote a paper on the effects of CH4 on the habitable zone. CH4 actually has a cooling effect for HZ planets orbiting cooler stars!
https://iopscience.iop.org/article/10.3847/1538-4357/aab8fa/meta
Thankyou for the update. My cursory look into this was that teh M-O approach was an empirical observation and that other mechanisms were adding to, but not entirely, superseding this explanation.
From purely the consciousness angle, it is my understanding we still have no clear idea what it even is, although fMRI studies with anesthesia seemed to have located the brain regions that were last activated before consciousness was lost. It remains one of the most argued issues in artificial intelligence and neural substrates.
If anesthesia was entirely mediated by the effect of gases on ion channels, then it seems likely that life on other worlds could easily evolve the proteins to deal with high nitrogen and carbon dioxide pressures. It might well be possible to design such proteins and test them in gene-engineered mice. [However, I am skeptical the M-O correlation is coincidental. Lipid solubility seems to be playing some part in anesthesia. Perhaps we can communicate about this offline after I have done some more research.]
“However, I am skeptical the M-O correlation is coincidental. Lipid solubility seems to be playing some part in anesthesia.Perhaps we can communicate about this offline after I have done some more research.”
Hi Alex. Yeah, the results of my own research yielded that even though lipid solubility may not be the entire story regarding anesthetic action, it is almost certainly a good part of it… The other realization was that there really isn’t another better alternative theory at this point, at least one that is as complete. I’d be glad to discuss this further offline if you want.
I am one of those clinging to the M-O hypothesis, as well. It just makes sense that dissolving a gas in a membrane would slightly change its physical characteristics, and that nerve conduction, especially the speed of propagation of the action potential, would be affected by this. It also makes sense that slowing nerve impulses enough would suspend the autonomous brain activity that constitutes consciousness. I think the observation held up against M-O can be explained without stipulating specific binding to proteins.
That said, there probably are modern anesthetics more complex than NO2 or chloroform that do have specific action. But, that is really beside the point.
“I have not had access to the full paper”…
Would this be it?
Yes, that’s it. My paper is open access, so it should be available to everyone.
Our best test case for the HZ and the CLHZ concepts is still Mars. Our lack of getting to the basic question of whether any life at all exists under the surface of Mars with probes suited to the question (or more obviously manned exploration to provide more flexible and more intelligent means of inquiry) over the last forty five years or more is disturbing to say the least. Some of the more powerful countries have cooperated to build a space station at a cost of more than 100 billion USD but we haven’t sent a single manned mission to Mars. I don’t understand this priority system. Single nations (even the US) would find it financially onerous to fund the entire cost of such missions but a consortium of nations could do it. Obviously we have at least two major concurrent crises to deal with: the pandemic in the short term, and human induced climate change in the longer term. Solving these issues and looking for life in the solar system should be achievable goals if we return to rational thought and science based investigation to drive our governance. Vote for change (you have a perfect opportunity in November in the US).
A space station was a more immediately practical task. It was also desired to sop up Russia’s expertise for political reasons at the time. A Mars mission still has not been architected, and we still don’t really know if we can build an ECLSS or whether supplies will have to be shipped for the full length of the voyage. The ISS has proven a huge money sink for rather limited gains. It will be interesting to see whether the US builds another space station after the ISS is decommissioned. I suspect not and China will be the only nation with an active space station. Perhaps the Russians will build another of their own.
The question of the value of humans on Mars still seems contentious. It is obviously cheaper to send robots. They also last a long time. Humans can certainly do things a lot faster, but the cost is huge. It may be mistake to think teh US could even lead in an international Mars mission. The Chinese might prove more aggressive in this regard on their own. Maybe SpaceX will be first as they seem to intend, but it would still be very expensive to support unless they can get some sort of external funding. As for science and seeking out life, I don’t believe that is a priority for them. Musk seems to have similar thoughts to Zubrin, which means that finding life on Mars is teh last thing either of them want as this would upset colonization plans.
With respect, we have put many robotic probes on Mars and they have produced many discoveries especially with respect to previous conditions such as a warm, wet early climate. However there has been no success (other than possibly one of the early Viking experiments) in detecting life or its metabolic products. A lab with humans to run the experiments and begin analysis of the data produced would surely revolutionize our knowledge about possible life forms. We will have to look underground and in the right places (near a water source is an obvious example whether frozen on the surface or possibly liquid below). I’m much less sure about the whole prospect of colonizing Mars. It could take many, many years to happen, even with the drive and determination of Musk and Zubrin. I would like to see some kind of real attempt to find out whether Mars harbours unicellular or even multicellular life in the next twenty years or so (strictly a personal wish based on my own life expectancy ha!).
NASA may *buy* their next, post-ISS station…from Bigelow Aerospace http://www.bigelowaerospace.com . (Their “test module” on the ISS, plus their free-flying Genesis I and II test modules in orbit, were developed with NASA’s–and other government space agencies’ and private companies’ needs–in mind.)
Narcosis is due to alterations in nerve conductivity by Membrane dissolved gases. Nerve conductivity is just one of many different mechanisms needed for animals, which in turn are only one segment of “complex” life. Some complex life (eg plants) does not even have nerves, at all.
Furthermore, narcosis is really only due to deviations from conditions an organism is adapted to. There is no reason an organism could not evolve a slightly different form of nerve conduction that is, for example, tolerant of ultra-high concentration s of chloroform, or whatever other gases you want to consider.
To use narcosis as a criterion applicable to life in general is, in my view, extremely farfetched. It should not impact our choice of where to look.
Wonderful to see you here again, Eniac. Was thinking about you and your many contributions just the other day.
Appreciate the kind words, Paul! I have been busy, but hope to be following more closely again going forward!
Hi Eniac,
The key assumption of the paper is that complex life (i.e., air-breathing animals, plants are not considered here) on alien worlds might exhibit similar respiratory limits as does complex life on Earth. Hence, the narcosis criteria is based on our current understanding of Earth animal biology. I do not think that this is far-fetched at all because we only have any idea of how animal life on this planet works. By extension, we also only have any idea of how carbon-based chemistry, water, and DNA-based life work. Such information forms the basis of virtually all operable HZ definitions anyway.
Plus, Earth is the only proven example of complex life in the universe. Other scenarios are much more speculative, as there is no way of currently knowing how far complex life can deviate from the Earth example on other planets. Even so, as I show in the paper, the CLHZ does get somewhat wider if it is possible to adapt to narcosis.
Now, simple life is indeed a different story and in our experience seems to be able to adapt to a wider set of conditions than can complex animal life. However, HZ definitions for that type of life already exist.
Simpler life forms when given multiple paths of evolution, might reach dominance in the paths that have competitive advntages. Exclusion of those paths and their competitive advantage may still leave open the other paths. The results further down various paths could even be mutually inconceivable.
I agree with all that, and a CLHZ is a good idea. I would just like to see it based on something more fundamental than narcosis. If I were to ask myself the question “why would complex life be prohibited by a high CO2 concentration, I would start by looking at respiration, which at least is common to all complex life on Earth, not just animals.
Just because I am uncomfortable with that one premise doesn’t mean I don’t like this work. It is well motivated and executed.
Hi Eniac,
No worries! Thanks for the comments. Constructive critiques are always welcome!
This work is concerned with respiration. It is just that narcosis is considered to be the baseline end state once that starts to fail. For instance, before CO2 narcosis becomes a major factor, renal composition helps counter the increase in blood acidity. Narcosis is only reached when this mechanism gets outstripped.
Here’s a recent article headline about anesthesia: “Anesthesia’s effect on consciousness solved, settling century-old scientific debate”
https://www.sciencedaily.com/releases/2020/05/200529150619.htm
An anesthetic like chloroform would change cell membrane lipid rafts from ordered to disordered, suppressing certain neurons from firing.
The change of state of lipids between ordered and disordered sounds very similar to what happens in liquid crystal displays.
A rather elegant solution if correct.
Almost certainly correct. Sounds like hype at first glance, but the story holds up under scrutiny. Lipid rafts and other order/disorder phenomena in lipids, proteins, RNA and even DNA are one of the few remaining “white areas” in the map of life that are currently being explored heavily. It is fitting that the long unsolved mystery of anesthetics turns out to have been hidden there.
Thanks for pointing it out, Randy Chung!
Looking at Dimetrodon from the Permian Triassic, I assume like animals today, it breathed an atmospheric pressure of only on bar, the atmospheric pressure has remained relatively the same since the beginning of the Cambrian explosion. I base this on volcanism which does not add that much to our atmospheric pressure.
Nitrogen narcosis occurs at about 100 feet. 33 feet is equal to on Earth atmosphere, so 100 feet under the sea is equal to three Earth atmospheres. I don’t think land animals had the problem of nitrogen narcosis because they would not be able to survive and thrive. When life or fish had to move from the sea to the land, a new adaptation was required based on necessity to a completely different type of environment to move around which included gravity and the need for legs or limb growth to walk on land through mutation without buoyancy and bladders to help them. Lungs also became a necessity.
There is the idea that Earth started out with a thicker atmosphere as much as five bars and it lost it through chemical reactions with the rocks, weathing and combining with sea water. There was also a collision with Theia where the Earth lost it’s atmosphere, but it was replenished from the release of water vapor and gases from the mantle. I like the model of thicker atmosphere in Earth’s past. The Sun was less luminous in the past. Consequently, Earth could retain a thicker atmosphere in its early history. The atmospheric pressure is mostly controlled by the mass and gravity of the planet so the larger Earth twins and super Earths are going to retain a larger atmosphere and pressure over time not including solar wind stripping. A larger gravity will also be a problem from animals which is why I don’t think the super Earths have any animals.
I also like a model where the atmospheric pressure has stayed relatively the same for billions of years. If this is general principle so it is contained in the Gaia hypothesis, then all exact Earth twins with a Moon will have these same environmental conditions.
This might well be a conclusion of the paper.
But could a super-Earth have plants, fungi, and possibly animals without a brain? Could any remote observation of biosignatures indicate whether a super-Earth had only plant life? Could a super-Earth have a dense atmosphere that was mostly H/He and very little N2 and CO2? (N2 is oxidized to NO2 and dissolves in the ocean, and there is no denitrification. CO2 dissolves in the ocean, precipitates out as carbonate, but very low volcanic emissions do not increase the CO2 pressure, which in turn is kept low by plant carbon fixation, IOW the geologic carbon cycle is not operating as it does on Earth.)
Then again. different biologies may allow animals to live in dense N2/CO2 atmospheres. We can only speculate based on our sample of one.
Oceans negate the higher gravity of super earths.
Super Earth’s have surface gravity of at most 2-3 g. All that would do is somewhat reduce the size of the largest possible animal, because the effect of gravity scales with size by some weak power law (2/3 if I remember correctly).
Alex,
Thank you for this excellent article. In these intriguing discussions regarding complex and/or simple life existing elsewhere in the Cosmos, I wonder if both might be rare for a similar reason.
Traditionally, there have been two main theories regarding the origin of life. One is the “genetics first” theory whereby RNA came before metabolism and the other other is the “metabolism first” theory whereby metabolic networks arose prior to RNA. To your knowledge, has anyone considered that a possible reason for why abiogenesis might be exceedingly rare is that a primitive metabolic network just happened to link up with genetic material (RNA or a pre-RNA molecule) perhaps within a micelle-like protocellular structure? The analogy that comes to mind here is that some think multicellular life is rare because it may have relied on an unlikely event such as an anaerobic bacterium swallowing an aerobic bacterium (aka The Endosymbiotic Theory)…
That rare event happened twice – once for mitochondria, and once for chloroplasts. However, it is more complicated than that, as the key difference between prokaryotes and eukaryotes is the separate nucleus containing the DNA. In prokaryotes, the DNA is not confined.
As for how life started, and whether either of those 2 models is correct, we just do not know. Unless we can test the hypotheses in the lab (wet chemistry or computer simulation), it might have to wait until the far future when survey craft can sample exoplanets to look for clues. Suppose those surveys found abundant life, but all of it looked like Earth-life – same molecular biology and basic metabolisms? Then won’t that imply a strong convergence to the only viable biology, or a common origin?
A slightly aging (2005) book on abiogenesis is Robert Hazen’s Gen.e.sis that I can recommend for a flavor of ideas and work. Progress has been slow and I haven’t seen any breakthroughs in this area. Maybe there cannot be.
Multicellularity is not rare as it has independently emerged many times. There is no such thing as a last common multicellular ancestor, because there were too many. Even metazoa or comparatively complex life forms have evolved at least twice, and the early origin of such complex life was delayed by oxygen level rather than by evolution of complexity itself. The first time was 2.1 Ga Francevillian biota, following Lomagundi oxygen overshoot. The second time was 0.6 Ga Ediacaran and Cambrian explosion, following Neoproterozoic oxygenation.
However, abiogenesis might be a rare event, since all life on Earth likely have a single last common ancestor, or all the extant life came from a single abiogenesis. This either means that abiogenesis only happened once, or the life from other abiogenesis all went extinct and left no descendents.
Actually, Earth’s last universal common ancestor (LUCA) is far younger than abiogenesis. It already had DNA, ribosomes, a genetic code, the works. Unfortunately, there is no way to use comparative genomics to look further back in time, so the era before LUCA is completely open to speculation.
However, since RNA is itself a product of metabolism, the RNA-first theory just doesn’t make sense. Metabolism is the only thing that is REALLY needed for life, so it must have evolved, first.
Ironically, there is some interesting work that indicates CO as the central driver for first metabolism, as it can provide both carbon and energy at the same time, the two most fundamental ingredients to living organisms.
Roughly, it works like this: CO + H2O -> organics(CHO) + CO2 + energy. Of course, you have to fit in nitrogen, probably from cyanide, and catalysts (nickel and iron, which are still found in central metabolic enzymes today), but the principle is pretty clear and compelling. CO and cyanide both occur in volcanic gases, to make things even more interesting….
This is a good start to investigate the CO theory of early metabolism: https://science.sciencemag.org/content/276/5310/245?ijkey=68e52357248309190315850e7786c37488ade6d8&keytype2=tf_ipsecsha
Another interesting read involving CO, cyanide, and iron in early life: https://www.nasa.gov/press-release/goddard/2019/cyanide-in-meteorites
Fascinating that there are inorganic complexes that resemble the active site of some of the most ancient metabolic enzymes, the hydrogenases. Presumably there are terrestrial occurrences of these minerals, such as at undersea volcanic vents, the most likely location where life might have started.
The subject of this article is an excellent argument for dispatching interstellar probes of at least the slow system fly-through variety, or–even better–the stellar rendezvous & planetary orbit type, with or without one or more landers for investigating each target system’s exoplanet(s) that appear(s) the most promising for indigenous life. Debates like the one in this article *are* helpful–they serve to help narrow down the list of exploration targets–BUT:
As with the 1950s and 1960s debate about whether the lunar surface was covered with a very deep layer of dust in which a landing spacecraft would be swallowed up as if it were descending onto quicksand (the “thin-dust” advocates’ arguments ultimately seemed the most likely to be correct, but only the first Luna and Surveyor hard landings [as opposed to the earlier Lunas’ and the Rangers’ self-destructive, lunar terminal velocity impacts] and soft landings demonstrated their correctness), only exploration with on-site instruments, in orbit and/or on the ground (perhaps around many stars) can settle the habitable zone width question, and:
This could be the case even if one or more of the probe-visited exoplanets was/were inhabited by complex–and even intelligent–life; as Arthur C. Clarke pointed out in his 1968 non-fiction book “The Promise of Space”:
“This proxy exploration of the universe is certainly one way in which it would be possible to gain knowledge of star systems which lacked garrulous, radio-equipped inhabitants; it might be the only way.” In addition:
Freeman J. Dyson also pointed out that intelligence does not and need not imply a technological civilization, as he was quoted (as the frontispiece of Chapter Seven of Ronald N. Bracewell’s 1974 and 1976 book “The Galactic Club: Intelligent Life in Outer Space”):
“I make a sharp distinction between intelligence and technology. It is easy to imagine a highly intelligent society with no particular interest in technology.” (Indeed, although he was not against technology per se, Dyson considered modern, high technology–as it is implemented in most human societies on the Earth–to be largely “a purposeless cancer of exploitation” [including the planned obsolescence of products, forever–and frequently–having to upgrade computers’ Operating Systems, and countless mathoms–a Scottish term for useless objects–that are produced and marketed to do things that human beings were and are perfectly able to do by themselves, etc.].) He feared that an extraterrestrial civilization–especially an older, technological one–that we might meet could be like humanity with respect to technology, but far more so, to our eventual detriment. As well:
If one of our exoplanet-orbiting starprobes spotted a happy, contented, but non-technological civilization, perhaps the best thing we could do for them would be to simply leave them alone, observing them from orbit for as long as the probe lasted (finally disposing of it in an ocean or an isolated land area, such as a polar cap or an active lava field), but without ever attempting contact, even “just” via durable, landed artifacts. Plus:
Ironically, leaving one or more artifacts on a moon, or elsewhere in their system–which they could only discover and understand after–and IF–they later developed spaceflight by themselves, much as with the Monolith in “2001: A Space Odyssey”–might be the best way to facilitate eventual contact, with them taking the initiative to do so. This would require patience on our part (a good thing!). It would also have a salutary effect on ^our^ society (“We need to stick around, and with modern cultures, so that ‘they’ will have us to talk and visit with [we could send Bracewell interstellar messenger probes–“electronic ambassadors”–and/or starships to visit them, if they signaled us by radio or laser after deciphering the artifact(s) left by the initial starprobes], if/when they develop the needed understanding, science, and technology to do it”).
Remote sensing will gain us a lot before we dispatch interstellar probes. Imaging planets, detecting various biosignatures, detecting plant life on continents, and other modalities will suffice to indicate the state of worlds in the HZ, CLHZ.
Orbiters around exoplanets will be able to image large animals, herds of animals, even possibly swarms of insects. Changes in atmospheric gases and changes in the landscape with seasons.
A lander[s] will be able to catalog the planet’s organisms and tell us a lot about their biology. Detailed observations will tell us about their ecosystems and the planetary biosphere. [The recent water sampling of Loch Ness DNA was able to detect a range of known organisms including fish and mammals, but no sign of Nessie.]
For me, the issue is, “where do we start to put our resources?”. Within the next century, assuming we continue to catalog exoplanets and model their likely surface conditions, we will have a very large number of candidate worlds to visit. Obviously, the closer they are, the more interesting they are to target using the known propulsion methods we have. [But let’s no rule out possible other means to visit very distant worlds in a timely fashion.] Proxima’s worlds would make a good test of interstellar probes due to its relative proximity. I suspect that the nearest world that has a good biosignature will prove much more distant. If the choice is a world that has only microbes versus one that looks like it has complex life but is more distant, that latter might be a more rational target.
Should the cost of sending interstellar probes drastically decline, then we should send them out to all teh nearer stars with planets, starting with stars with worlds in the HZ. We will want to catalog them in detail, especially those with life. These probes need not be all or nothing scientific platforms. First send out tiny, minimal craft to make a quick assessment of such worlds from a flyby. If the data look interesting, send increasingly sophistical probes. This is a very long term project after all, worthy of a long-lived, technological and curious, civilization. Whether we are such a civilization, I don’t know. IMO, the jury is still out on that.
I agree–like Bracewell, I see starprobes and SETI/exobiology (in the broadest, most inclusive sense, encompassing remote detection of life-supporting conditions and life itself, even unintelligent life) as complementary approaches to achieving the same goal, not as adversaries. Something that is happening *right now* will make both methods–“going there, and scrutinizing from afar (here)”–less difficult and expensive:
NASA, ESA, the space agencies of China, Israel, Russia, and India, as well as several private U.S. and Israeli aerospace companies–“new space” as well as legacy firms–are all engaged in a “frenemy”-type race to reach the Moon, establish bases there, and industrially (as well as scientifically) utilize both the ^physical^ lunar resources and our natural satellite’s “phenomenological resources,” and:
These include–as Neil P. Ruzic covered (their many uses) in his 1970 book “Where the Winds Sleep: Man’s Future on the Moon–A Projected History” (his 1965 book “The Case for Going to the Moon,” which he wrote to try to get the scientific community excited about Apollo’s potential, is also well worth reading)–the free, hard vacuum and cryogenic cold, which are available in unlimited quantities on the Moon, along with readily, locally-accessible metals, minerals, and even water. Other such “phenomenological resources” of our Moon include:
[1] The two-week (to effectively eternal, in permanently-shadowed craters) periods for taking very long astronomical photographic–or CCD–image exposures;
[2] Very precise determination of the coordinates of optically invisible objects (by using crater rims [along with local atomic clocks] as part of crater bottom-mounted X-ray and gamma ray telescope systems), and;
[3] The electromagnetic shielding provided by 2,000+ miles of solid rock, enabling lunar farside radio telescopes to receive even the faintest signals from galactic and extragalactic sources–and, perhaps–purposely-directed or incidental intelligent signals (between stars, between stars and interstellar vessels, planetary radar pulses, etc.) from other civilizations. Now:
Building up a lunar transportation infrastructure, bases (Elon Musk has recently proposed a base, intended to eventually become a permanent settlement [rotating buildings in lava tubes could provide 1 g and radiation shielding], which he called–what else?–Moon Base Alpha…), lunar mining and industries, and local scientific stations, would facilitate both high-resolution observations of exoplanets and the production, launch, and operation of interstellar space probes. In addition:
Optical, radio, and other spectrum-region telescopes of unprecedented size and sensitivity–yet of much lower mass and greater physical simplicity, thanks to the much weaker lunar gravity, lack of atmosphere (and ^its^ resulting absence of sky light, making observations possible even when the Sun is in the lunar sky)–make the Moon an ideal location to set up telescopes (and other instruments) to, like H.G. Wells’ Martians, “scrutinize and study, closely, the surfaces–and any living things thereon, or on the seas or in the skies–of the planets of other stars.” As well:
The same lunar development will also make possible the local production, launch, acceleration, control, and data & images reception from interstellar probes. Laser light-sail probes of the type that the late Dr. Robert Forward designed (all spin-rigidized disc sails; single-stage, stellar system fly-through probes and two-stage, stellar rendezvous probes that could explore their entire target systems, using the local star’s [or stars’] light to operate as solar sails) could be manufactured on the Moon, and be launched and accelerated to significant fractions of the speed of light, pushed by Moon-based launching lasers, and:
Dr. Forward’s laser light-sail probe designs were rather large (his 3.6 kilometer-diameter, single-stage, one-metric ton [about one-third each of payload, supporting structure, and sail panels] Alpha Centauri system fly-through probe would reach 0.11 c, pushed at 1/30 g for three years by a 65 gigawatt Sun-pumped laser array).
His two-stage, 0.21 c peak velocity stellar rendezvous probe design (to be pushed at 0.005 g by a 7.2 terawatt laser array, reaching the Alpha Centauri system 41 years after launch) was even larger and heavier (785 metric tonnes in total)–its sail would be 100 kilometers across (consisting of a 30-kilometer central disc sail and probe massing 71 tonnes, including a 26-ton science payload). The outer ring sail deceleration stage would separate near the target stellar system, and the central 30-kilometer rendezvous sail would turn around 180° to face it, so that the “bundled” laser beam from the Solar System would slow the rendezvous sail–which would henceforth utilize the local starlight to operate as a solar sail, to explore the system’s planets–to a stop in the Alpha Centauri system. Both laser light-sail probe designs would have required a 1,000 kilometer diameter, 560,000 metric ton flat plastic Fresnel-zone-plate lens, free-falling very slowly (or levitated) between Saturn and Uranus, to collimate the laser beam (for interplanetary light-sail ships, the Fresnel lens wouldn’t be needed, and their propulsive lasers could be based in Earth orbit, instead of at the planet Mercury). BUT:
Technological developments since then (the 1970s – 1990s) have reduced the practical payload masses–for both the stellar system fly-through and the stellar rendezvous laser-pushed light-sail interstellar probe types–to less than ten kilograms to a few hundred kilograms. Such smaller and lighter payloads could be pushed by smaller and lighter sails, which would in turn be able to utilize smaller, less powerful, easier to build, and cheaper laser arrays, and:
Such laser arrays–which could also be used commercially, to push interplanetary light-sail ships going to and from high Earth orbits, the Moon, planets, moons, asteroids, and comets–could be built on the Moon, with the laser cannons (which could also be Moon-built) set up either at both lunar poles (the permanently-sunlit peaks [“Peaks of Eternal Light”] in those places could provide continuous solar-electric power for the lasers), and/or evenly-spaced along two circles of longitude girdling the Moon, with the circles 90° apart (to enable the computer-linked lasers to provide continuous illumination of, and thrust on, the interplanetary and interstellar light-sails; the non-polar lasers could be nuclear powered, or powered via power lines from distant power plants). Plus:
Instead of one “bundled” beam, the multiple laser projectors–at the lunar poles, and/or spaced around the longitude circles–would be controlled and aimed so that their multiple beams intersected on the vehicles’ sails (lower-power guide laser beams from the Moon, and/or pilot beams from the light-sail craft, would facilitate accurate “thrust laser beam” aiming, and accurate placement of the craft within the beams’ intersection zones). Moon-based telescopes could be used to select the probes’ targets, thus saving the exobiology program time and money overall.
HZ is almost certainly narrower than what is widely adopted in literature, without even considering the potential intoxication from atmospheric composition.
In order to evolve complex life, surface liquid water must be stable on geologic timescale, or billions of years.
The planetary system must be able to cope with instantaneous releases of amount of carbon that is several times more massive than all the fossil fuel reservoirs and resources in the crust, such PETM, PTME, or CPE events. As it has been hypothesized that LIPs might have desiccated ancient Venus (Way & Del Genio, 2020), that is to say slowly rotating HZ inner edge is ineffective against LIP eruptions and could be extremely susceptible to desiccation. In fact, Popp (2016) 3D simulation has shown that with only 3% increase in solar luminosity, LIP eruption could already permanently put Earth in moist-greenhosue state, much closer than classical inner edge.
The planetary feedbacks must be also effective against permanent global glaciation. HZ studies such as Kopparapu (2013) has always assumed “warm start”, ignoring the fact that planets inevitably enter glaciation at some point. The feedbacks to effectively deglaciate the planet seem only to work when insolation is above 0.45 (Turbet et al., 2017; Kadoya & Tajika 2019)
What’s more, HZ calculation also assumed ideal C-S cycle. But it might not be the case given how sensitive C-S cycle is to soil lithology, orogeny, land fraction and hydrology (Graham & Pierrehumbert, 2020). Perhaps Earth being fine-tuned is possible.
These may give a glimpse into hemoglobin dissociation curves marine vertebrates. And also of marine hemocyanin dissociation curves.
And marine hemerythrin dissociation curves.
And an offering of images:
Marine hemoglobin dissociation curves
Marine hemocyanin dissociation curves
Marine hemo erythrin dissociation curves
Alex Tolley, a lack of a carbon cycle will result in a larger greenhouse effect since there would be no plate tectonics and no subduction zones to subduct old crust and melt it and rifts where new crust is being formed and spread out. The larger mass super Earths have been postulated to potentially have that problem. Could large ocean worlds still have sea animals like large whales, dinosaurs etc? The atmospheric pressure could make a difference in surface temperature. A five bar atmosphere will still have a big greenhouse with no carbon dioxide. A three bar atmosphere will have a more efficient greenhouse effect than a one bar atmosphere.
A lack of biosignatures like oxygen and methane would rule out animals and sea creatures.
I agree that on a lifeless super-earth, especially a water world, the geologic carbon cycle would not work to stabilize the CO2 in the atmosphere. But call that the atmospheric pressure is only affecting complex life, and possibly only metazoa. CO2 will dissolve in water making it more acidic, but this may be taken up by microorganisms. Photosynthesizers will fix that carbon, and shelled organisms with convert it to carbonates. Both of these processes could draw down the CO2 and deposit it as rock and organics on the ocean floors, layer by layer. Sedimentary rock doesn’t need subduction by plate tectonics. If that could work, then a dense CO2 atmosphere could be drawn down to low pressures, just as it has on Earth. The nitrogen is more problematic, but if there is abundant oxygen from those photosynthesizers, then it will oxidize and dissolve in the oceans. It will remain there until denitrification occurs, which may not occur unless the appropriate microorganisms are present in the oceans. So I would hypothesize that dense nitrogen atmospheres could be depleted too.
Purely speculation on my part as a possible way to reduce atmospheric pressures if needed to make a more suitable world for life. An exo-Gaia hypothesis for such worlds, if you will. However, if abiogenesis is not possible on such super-Earths, then this cannot happen. Having said that, haven’t there been suggestions that some super-earths may be more supportive of life than Earth?
I agree that super Earths may well be more suitable for life. I see no reason why a thick atmosphere should be a hinderance, and gravity certainly doesn’t make a difference except for the very largest land animals. If a thick atmosphere makes the world too hot, just pick another world a little further out where it’s cooler. In other words, we may have to move the HZ outwards a little bit for larger planets, but that would not impact the number of “habitable” planets we would expect to find of that size.
As to abiogenesis, we really have no idea what makes it possible/impossible. Currently the most attractive place (imho) for it to have happened is deep sea vents. There is very little on the surface that impacts those environments. In particular, the presence or absence of an atmosphere would not make a difference. Neither would intense radiation of any sort. A world like Europa in the outer system may well qualify, for example. I don’t think there is good reason to believe that super-Earths would have less or different deep sea vents, so large planets way outside the HZ may well be suitable for abiogenesis. Maybe more so than Earth, because the outer system has way more water.
It is important to realize that both photosynthesis and respiration are comparatively modern developments on Earth. Early life would have resembled the chemotrophs we find under anoxic conditions today, unable to exploit the energy of light and looking at oxygen as a dangerous poison. Gravity would not have made the slightest difference to them. If, as most believe, early life was submarine, neither would atmospheric pressure. And “early” here really means “most of the time, except for the most recent past”.
Had just put together a comment and was about to submit when the phone rang. Think I lost it. But to recap: This is a very interesting discussion from the standpoint of biochemistry. Thanks for formulation and comments. Saying that, came away with two contradictory impressions about defining /refining the habitable zone:
1. Scattershot vs. narrow scope? I had to imagine a farmer confronted with animals foraging in a cultivated field at night. ,,
2. On the other hand, I am inclined to think that the HZ narrows
with temperature more than figure 2 suggests. If we take the main sequence lifetime of the sun as a reference point of about ten billion years, then the brightness of other stars is telling about the duration for HZs. It might be over with before a planet gets to form or cool.
Given that the mass luminosity relation is of a power between the third or fourth, then low balling the case for a star ten times more massive than the zone, the main sequence duration would be about ten million years. My understanding is that circum-stellar disks remain stable for only about that long themselves. HZ planets would be in big trouble even before they had fully formed.
Of course, stars of ten solar masses would be considerably hotter than 9000 Kelvin. The issue might be more pertinent to A and F stars with masses – or stars with masses two or three times that of the sun.
Assuming 3.5 power law for a 3 solar mass star, the lifetime would be reduced to 1/47th of the ten billion years, about 215 million. That is
not much geological history. I would have to wonder if a terrestrial planet would have sufficient time to cool. “Biogenesis” would need to originate from elsewhere.
We have often discussed G, K and M stars, perhaps on account of such concerns being implicit. Since more massive MS stars do have larger disks, there might be a little additional difficulty in detecting terrestrial stars orbiting them, but I wonder if there has been much examination of their numbers in brighter stars, say, in the Kepler data base.
The conditions for life around G class super Earths in the life belt will be good early in the life of the star where the greenhouse counter the lower luminosity. The problem is by 4.5 billion years will the planet be too hot. The reduction of the atmosphere will be harder for life with a larger escape velocity and gravity than Earth. The denitrifiers or nitrosomonas and nitrobacter convert the nitrates into nitrogen gas or molecular nitrogen. They put the nitrogen back into the atmosphere which makes more atmosphere and are part of the nitrogen cycle. How much life has contributed to the total weight of our atmosphere during the history of life with nitrogen I don’t know.
One hopes with the super Earths that the temperatures are not too high from the start or early on. It might be different for a super Earth which is just outside the life belt at 4.5 billion years which would have a cooler atmosphere, but enough greenhouse effect to support life?
The spectra will tell us.
Molecular oxygen is two oxygen atoms joined together, and on Earth it is essential for life. Comet 67P/Churyumov-Gerasimenko has abundant O2, so comets may have increased these levels throughout earth’s history. When a star passes close to our sun it disrupts the Oort Cloud and sends many large comets into the inner solar system that break up and impact earth. These large comets increase O2 levels and may have been essential in the development of advanced life.
This also happens much more frequently in the small M Dwarf planetary systems, which would keep O2 levels high and aid in the development of intelligent life. This would also expand the Habitable Zones for complex life around them.
Molecular oxygen in comet’s atmosphere not created on its surface.
https://phys.org/news/2018-07-molecular-oxygen-comet-atmosphere-surface.html
Alex Tolley, I would like to have your comments and criticisms of this train of thought. Tu.
Assuming cometary impacts were fairly constant over time, it would suggest that O2 levels would continue to increase, or that oxidized compounds would increase. We know that this does not seem to be the case in teh deep past, and today the O2 levels respond to seasonal plant growth (CO2 drops with teh Northern summers and the O2 slightly rises). Any addition from comets would be marginal at best.
In the deep past, free oxygen was created by photolysis of water. It is possible some of that water came from cometary impacts. For comets to change the O2 level the mass from impacts would have to be on a scale that was a significant fraction of these known processes. I am not aware of any data suggesting this is teh case. Do you have any references to indicate otherwise?
I agree with Alex. Also, I believe the O2 they talk about is extremely rarefied. The “atmosphere” of a comet would be considered a perfect vacuum on Earth, and very little of it is oxygen.
The development of advanced life is clearly linked to the advent of photosynthesis, which is both necessary and sufficient to explain the subsequent oxygenation of the Earth.
Read the article, that is what they are talking about.
Co-author Dr. Marina Galand, from the Department of Physics at Imperial and Science Co-Investigator of the Rosetta Plasma Consortium, added: “Surface generation of molecular oxygen may still happen on 67P, but the majority of the molecular oxygen in the coma is not produced through such a process.”
The new analysis is consistent with team’s original conclusion, that molecular oxygen is most likely primordial. Other theories have been proposed, and can’t yet be ruled out, but the primordial theory currently fits the data best.
“This is also supported by recent theories which revisited the formation of the molecular oxygen in dark clouds and the presence of molecular oxygen in the early Solar System. In this model, molecular oxygen created froze onto small dust grains. These grains collected more material, eventually building up the comet and locking the oxygen in the nucleus.”
Yes, photosynthesis, is what makes it now but CO2 was the primary atmosphere before. Historically most of the earth’s crust was destroyed in the iceball earth some 700 million years ago so any older iron sinks were probably destroyed along with any advance lifeforms. The oceans records are no more then 270 million years old and most much younger because of subduction. There are reports of older periods of oxidation but most of that history has been destroyed, CO2 was probably where most of the O2 ended up along with iron sinks.
Cometary impacts are not constant over time but peak when the solar system goes thru the denser galaxy arms. This also increases our encounters with other stars entering the Oort Cloud and kicking them into the inner solar system. A lot more star encounters and dense dust clouds high in O2. The giant comets may come in waves for millions of years with earth impacts happening for 10,000’s of years from a comet breaking apart after passing close to the sun.
Three quarters of the earth’s ancient history is gone in the oceans because of tectonics and the majority of the earth land area has been erased by snowball earth with just a few of the large deep impacts still existing on the oldest crust. I think we need a good detective to put together what happened in the 84% of earth’s history that is now gone. Do not forget all those iron/nickel asteroid impacts leaving all the iron sump laying around.
The M dwarfs have a more congested airspace around them with the planets in the HZ traveling 33 times faster then earth. This would make for many more impacts and most of these system seem to be richer in lighter elements so more O2. For example the Trappist 1 system has five of its planet with a lower density that is closer to Mars and we know Mars has had liquid water on it.
I think you may be under a misapprehension about what is known geologically. While the current ocean floors do not go back that far, they have also become continental material that has. We have rocks that can be dated to nearly the age of Earth. We have rock samples with evidence of life at over 3.5 bya. Almost any paleo chart will go back over 3 billion years with proxy data from sediments that indicate atmosphere composition, including the changes as teh Great oxidation event proceeded. The Cambrian “explosion”when the huge diversity of phyla appears is about 540 mya.
My understanding is that water from asteroids and comets makes up some fraction of the water in the Earth. Any photolysis of that water would produce O2, at least transiently. We know that any free O2 generated by photolysis(which was small) was quickly used to oxidize reduced compounds. It was the evolution of photosynthesis and teh steady product of oxygen that lead to the Earth’s atmosphere accumulating O2 to the level it eventually became.
The paper you cite indicates that the O2:H2O ratio is about 1:100. IOW, the O2 contributed by comet impacts similarly composed would be around 1% of the water they delivered to Earth. I leave it up to you to show that this amount of O2 would be more important than the O2 produced by photolysis and that this would have shortened the time needed for photosynthesis to create an O2 atmosphere. I suggest you try some BoE calculations to see whether your idea would make a material difference to the history of the Earth. Then check it against what is known. For example, if the LHB was due entirely to comets, not asteroids, there should be a spike in that period of all the compounds in the comet, not just O2. Is there any evidence for that you can find? We do not know when exactly life appeared on Earth, but we do know that it was anaerobic. O2 would have been toxic. Would this have been more detrimental to emerging life than the physical LHB itself? Perhaps you can offer some falsifiable hypotheses about this early O2?
“I think you may be under a misapprehension about what is known geologically.” No, quite the opposite, I was a geology major at the University of Oregon in 1973. My favorite book on just what you are talking about and way ahead of its time “The Rediscovery of The Earth” 1979 english version, has been in my personal library for 40 years. (cost me $35.00 dollars) This book was talking about early life at 3.4 billion years and had 20 authors that were experts in their field, it was written in italian in 1975!
“Ancient rocks show high oxygen levels on Earth two billion years ago.” MAY 12, 2020
“Earth may have been far more oxygen-rich early in its history than previously thought, setting the stage for the evolution of complex life, according to new research by scientists at the University of Alberta and the University of Tartu in Estonia. The study provides evidence for elevated oxygen levels 2 billion years ago and flies in the face of previously accepted models.
The international team of researchers, led by UAlberta scientists, studied a Russian drill core containing shungite—a unique carbon-rich sedimentary rock deposited 2 billion years ago. The material provides several clues about oxygen concentrations on Earth’s surface at that time, including strikingly high levels of molybdenum, uranium, and rhenium, as well as elevated uranium isotope ratios.
“These trace metals are only thought to be common in Earth’s oceans and sediments when oxygen is abundant,” explained Kaarel Mänd, a Ph.D. candidate in the University of Alberta’s Department of Earth and Atmospheric Sciences and lead author of the study. “These trace metal concentrations are unrivaled in early Earth’s history, suggesting elevated levels of oxygen at the time when the shungite was deposited.”
What’s puzzling, Mänd explained, is that many widely accepted models of Earth’s carbon and oxygen cycles predict that shungite should have been deposited at a time of rapid decrease in oxygen levels.
“What we found contradicts the prevailing view,” says Mänd, who is completing his Ph.D. under the supervision of Professor Kurt Konhauser. “This will force the Earth science community to rethink what drove the carbon and oxygen cycles on the early Earth.”
The new findings also provide insight into the evolution of complex life. Earth’s “middle age” represents the backdrop for the appearance of eukaryotes. Eukaryotes are the precursors to all complex life, and require high oxygen levels in their environment to thrive. This study strengthens the idea that suitable conditions for the evolution of complex life on early Earth began much earlier than previously thought.
Future research will examine the delay between the initial rise of oxygen and the appearance and spread of eukaryotes, remaining an area of active research, one that University of Alberta and University of Tartu researchers are well positioned to help answer.”
This paper, “Paleoproterozoic oxygenated oceans following the Lomagundi-Jatuli Event,” was published in the journal Nature Geoscience.
https://phys.org/news/2020-05-ancient-high-oxygen-earth-billion.html
Two of the largest impact craters are also from the same period;
Vredefort crater.
https://en.wikipedia.org/wiki/Vredefort_crater
Lake Mistassini.
https://en.wikipedia.org/wiki/Lake_Mistassini
Late Heavy Bombardment Myth Is Dying.
https://crev.info/2018/01/late-heavy-bombardment-myth-dying/
The solar system travels thru the galaxies arms about ever 120 million years but if you look at the picture you can see that at times the sun will be longer in sections of the arms. There are also at least 2 to 3 smaller arms and the sun travels up and down the galactic plane as it orbits thru the arms. https://upload.wikimedia.org/wikipedia/commons/3/30/Galactic_longitude.JPG
These arms also are the place of birth and death of the
giant O and B type stars that explode as a supernova every million years. When they explode they pollute the arms with the volatiles elements from the nucleosynthesis.
https://upload.wikimedia.org/wikipedia/commons/thumb/3/31/Nucleosynthesis_periodic_table.svg/1000px-Nucleosynthesis_periodic_table.svg.png
As you can see in the chart, Oxygen is created in the supernova of massive O and B stars which live and die in the galaxies dense arms.
If I understand the Mand (2020) paper correctly, they argue that the Lomagundi-Jatuli Event (LJE. a large deposition of carbon in the sediments around 2 bya) indicates a much higher pO2 (perhaps 10+% pr present levels) that is generally accepted. Other papers do not confirm this analysis using other proxies such as chromium (Cr) ion ratios.
But let me accept for the sake of argument, that this event that precedes the rapid rise in O2 levels between 0.5 – 1 bya is correct. YI think you are arguing that:
1. This O2 may be non-biologic and come from primordial O2 from comets that were created by supernovae as teh solar system migrated within the milky way.
2. This event, and possibly others, may have influenced the course of the development of complex life.
As regards point 1, I would ask why we don’t see more of these events. Shouldn’t they be dotted around the stratigraphic record? On the face of it, the amount of O2 to account for the LJE would be huge, implying a lot of comet impacts at this time. If so, shouldn’t there be other evidence of such a spate of impacts occurring at this time? The paper indicates that the effect was transient, and that the O2 levels returned to the low levels seen on the proterozoic fairly quickly. Then we have over a 10^9 yrs of low O2 levels and then the appearance of complex life.
As regards point 2, whey the billion-year gap between teh LJE and the appearance of complex life. Doesn’t this decouple the 2 events? The creation of a temporary slightly more oxic condition in te oceans doesn’t seem to have done anything observable, and I am not sure what it could have done. The appearance of metazoa in teh Ediacaran period is coincident with the rise in oxygenation, and this leads to the Cambrian explosion around 540 mya. We see no such evidence in the sediments for complex life 2 bya. If the LJE caused a rise in O2 that stimulated complex life, wouldn’t we expect to find some evidence for this in the rocks such as traces, even of burrows or casts or tracks?
Constraints on Paleoproterozoic atmospheric oxygen levels
A case for low atmospheric oxygen levels during Earth’s middle history
A BoE calculation of the size of a comet to increase the O2 of Earth.
Current atmosphere mass = 5E18 kg
=> O2 mass 1E18 kg
Assume LJE event increases O2 to 10%, => 5E17 kg
Assume O2 = 1% of comet mass, comet = 5E19 kg
Assuming the comet is mostly water ice, using water density = 1 gm/cm3
=> 5E19 kg => 5E16 m^3 => 5E7 km^3 => diameter 450 km
For comparison, the largest comet is Hale-Bopp at about 100 km diameter.
So either is was a very large comet or a swarm of ~100 Hale-Bopp sized comets, or a far more O2 enriched comet/swarm.
Since you are a trained geologist, why do you say:
We have billions of years of ocean records in the sediments now part of continents. You must mean something different from what I understand as “destroyed”. When I think of destroyed geology, it is the first half-billion years of Earth’s formation that we have no record of anywhere in the crust.
Point 2 is arguable.
LJE is a transient event that did not result long-lasting oxygenated environment. We don’t really have a constrain on atmospheric pO2 during LJE but possibly 10% present-level. Earth soon deoxygenated around 2.0 – 2.1 Ga, and pO2 was below the threshold of metazoa until Neoproterozoic Oxygenation Event.
Fossils record is scarce but there is evidence to life forms comparatively complex to early Ediacaran animals emerged during LEJ. They are known Francevillian Biota. They too left traces and burrows so large that would be commonly assumed to be Phanerozoic animals (El Albani et 2019). They had size of a human hand, fed on microbial mats, grow in radial structure, etc (El Albani et 2010, El Albani et 2014).
But Francevillian biota disappeared immediately in the above deoxygenated strata, and such complex life did not appear again until Ediacaran.
I am very skeptical the Francevillian fossils are metazoa. While the fossil evidence has various interpretations, the genetic evidence we have suggests that the metazoa appeared much later during the Mesoproterozoic Era (1 – 1.6 bya).
So while it is not impossible some extinct metazoa appeared then, the genetic evidence of the life we know of does not seem to support this hypothesis. Maybe they are the long-awaited evidence of alien life? ;)
No way Francevillian biota could be any kind of animals as we know. I believe the common ancestor of extant animals is no earlier than 650 Ma based on molecular study (Cunningham et 2017). Even the common ancestor of extant eukaryotes is no earlier than 1850 Ma.
But in terms of complexity, they bear similarity to early Ediacaran animals.
If earthly in nature, Francevillian biota is probably some completely different complex life evolved during the transient LJE. It could represent another path to life form of high complexity, and possibly intelligence.
As to 1, I have explained in proceedings comments that the snowball earth destroyed much of stratigraphic record and both external comets and Oort cloud comets plus dust high in O2. Ocean impacts are gone for 95 percent of earths history. The ancient earth history in oceanic sediment is part of the giant jigsaw puzzle around the old continetal cores and much of it is metamorphic. Lets say for example that our current global warming creates a new snowball earth and humans destroyed themselves. A half billion years of 5 mile thick ice would wipe the earths surface of any trace of mankind and the oceans would corrode and digest any traces left in a billion years. The known impact craters from before 600 million years would also be gone.
The earth’s giant conveyer belt has recycled so much of earths history. We are still finding what happened before snowball earth and just how many snowball earths where there before the last one?
You make it sound like we just go out and drill and have a perfect core with 4.5 billion years of earths history instead of a spotty jumbeled mess that a forensic expert would be crazy from. I have given you what you ask for and explained why we still have a small amount of data on what happened for much of earths history.
This argument you present is a prime example of the problem of the academic communiy inability to except data that conflicts with their pet theories. Oh, answer to 1 explains 2!
While I am not a geologists, I do not accept your argument that the relevant geologic record is “wiped out”. TI see no way to argue that there are proxy data of this LJE period and top also say that the record in geology is missing. The Mand paper acknowledges that their proxy may be interpreted either way (high or low O2) but opt for the high O2 interpretation. Other proxies show no evidence of an toxic period at the LJE period. Therefore you are really left with the raw observation of a rich carbon layer that is interpreted as a result of increased deposition due to an more oxygenated event. This proxy for O2 as a % of (present atmospheric level (PAL) has a wide range. It is not a solid finding, but rather a controversial one.
However, as I said, if one accepts the argument, then it has to be explained. Your hypothesis is that it was delivered by a comet[s]. If so, it has to be a large mass of cometary material. It would have to be one massive comet that hit the ocean without breaking up to leave no other trace. A swarm would hit the continents too, leaving traces. However, comets are made up of a lot of other material and one might expect them to leave some other evidence in the record. An alternative hypothesis is that the event could be due to biological activity as photosynthesis had evolved over 3 bya.
To overthrow the multiple lines of evidence that supports the current view of life on Earth would need very strong evidence. Bear in mind even the Alvarez’s discovery of the iridium layer and the subsequent discovery of the Chicxulub meteor impact site doesn’t convince everyone that this meteor was the sole reason for the extinction of the dinosaurs (apart from the birds), and the meteor evidence is incontrovertible, as is the time period for their extinction.
Lay out a convincing story with solid evidence that does not contradict what we already know and I will be open to changing my mind. So far I haven’t seen it. If you are that convinced, why not write a paper and submit it to arxiv or biorxiv with a view to a possible peer-reviewed publication later?
Why not suggest to our host that you write a post about your hypothesis? It would be much less work than a paper and it should promote some lively discussion.
That’s why I had you scrutinize it to see what needed to be worked on plus it helps me fill in the details of the concept. Thank you and Nicky for your help and I may just do that but need to digest it and come up with more evidence. Could use some help with material on current research in ancient earth’s atmosphere and life, maybe a list of some of CD articles.
There is no evidence to support Lake Mistassini as a impact structure, other than the arc shape.
Vredefort impact has been dated to be 2023 ± 4 Ma (Jourdan et 2012), later than 2057 ± 1 Ma the termination of LJE (Martin et 2013), and rather coincident with productivity collapse at 2018 Ma and deoxygenation after LJE (Hodgskiss et 2019; Mänd et 2020).
“LHB is dying” only argues against the hypothesis that there was a spike in impact flux, which is likely true. It does not contradict with the overwhelming evidence of much higher impact flux in Hadean and Eoarchean.
The amount of oxygen delivered to surface by impactors is very insignificant compared to mantle degassing and sinks. Any sign of impact delivery is immediately erased by the long-term, balanced Earth system, because the sources and sinks are so huge compared to the little delivery by impactors. The stepwise oxygenation rather required a shift in Earth system as whole, such as mantle oxidation state, emergence of subaerial landmasses, increased continental weathering, etc.
I think this has become a one sided argument with a very major bias to impactors changing earth’s environment. So goodbye to the aforementioned dinosaurs!
Oh, and the Mastodons, they were such cute creatures to end up in such a mess! (YDIH 12900 years ago) https://cosmictusk.com/martin-sweatman-younger-dryas-2/
Please, stay safe, do not forget to wear your mask and hard hat when outside! ;-}
As you know, the theory of continental drift took a long time to be accepted despite the evidence. The KT impact hypothesis has a lot of evidence Evidence for impact.
What is the evidence for a cometary impact oxygenating the atmosphere and triggering the development of complex life? AFAICS, it starts with a contentious evidence of an increase in atmospheric O2 around the LJE. There is evidence that comets have a small fraction of O2 as part of their composition. But there appears to be no solid evidence of an impact or impact[s] as there is for the Chicxulub impact. The evidence that extant life was in some way driven towards complexity is based purely on some controversial fossils but there appears to be no corroborating evidence in the genetic evidence. This hypothesis strikes me as very thin at best.
Some things may be unknowable because there is no evidence. Is life on Earth due to a local abiogenesis? Were there multiple geneses and ours dominated driving the others extinct? Some things become discoverable by new lines of evidence. The idea that eukaryotes are the result of encapsulting other bacteria is now firmly established, despite it being initially rejected. Lynn Margulis also hypothesized horizontal gene tranfer in some organisms that resulted in the different life stages of many invertebrates. While this has not happened in humans, we now know that retroviruses are the source of some human genes. For any new theory, there needs to be evidence which explains the observations in a better way than an earlier theory. You know the various theories about earth’s gelogical history. In biology, Darwin’s idea of evolution by natural selection proved immensely powerful, sufficiently so to overturn the Biblical story of creation for all but the most diehard creationists. Evolution by natural selection has proven extremely robust with new lines of evidence that have supported the theory, not overturned it.
As I said, write up your theory laying out the evidence to support it. Ifit can resist being picked apart, you may have something.
The target rock that was excavated in the process of impact changed the environment in dinosaurs extinction, not the volatiles brought by impactor itself. The target rock of Chicxulub was 30% sulfate and 70% carbonate.
Even that change was only transient, Earth system also recovered immediately (million-year timescale) to the pre-impact state, and the extinction lasted only a few decades to a few centuries.
The causal link between impact and Younger Dryas is far from proven, see recent review by Daulton et (2017), Van Hoesel et (2014), Holliday et (2020). I would say it’s even in the state of lacking critical evidence.
Regardless, Earth system feedbacks mitigated Younger Dryas in millennium timescale and back to its previous trend.
Again, long-lasting, major changes such as oxygenation require fundamental shifts in Earth system.
Two major events, 500-800 Ma Neoproterzoic Oxygenation Event and 450-400 MaPaleozoic Oxygenation Event are rather coincident with a period of much less impact flux in Earth history. Impact flux increased after the end of the Paleozoic (250 Ma), but we don’t see rise of oxygen anymore.
Oldest known asteroid strike may have ended Snowball Earth.
More than 2 billion years ago, an asteroid slammed into Earth and created a 43-mile wide crater in Western Australia’s outback. Now, researchers believe it is the oldest known impact crater, predating others by 200 million years. And it’s possible that the impact led to a great thaw as Earth exited a global frozen state.
https://edition.cnn.com/2020/01/21/world/earth-oldest-asteroid-impact-scn/index.html
Precise radiometric age establishes Yarrabubba, Western Australia, as Earth’s oldest recognised meteorite impact structure
https://www.nature.com/articles/s41467-019-13985-7
Gene Shoemaker
https://www.famousscientists.org/gene-shoemaker/
I met him in 1989.
Again, even if it did change, it was not by the volatiles brought by the impactor itself, it was the impact target that undergoes melting had changed environment.
So it in no way supports atmospheric events are governed by impact flux.
Additionally, we don’t have evidence of casual link between deglaciation and Yarrabubba impact, other than the temporal constraint.
And again, two 500-800 Ma Neoproterzoic Oxygenation Event and 450-400 Ma Paleozoic Oxygenation Event occurred within a period of much less impact flux in Earth history. Impact flux increased after the end of the Paleozoic (250 Ma), but we don’t see rise of oxygen anymore.
Alex Tolley, most of the free oxygen was created by photosynthesis. The photolysis of water vapor occurs at high altitude in our atmosphere in the mesopause. Although this might potentially produce a false positive exoplanet spectral signature, I don’t think it is enough oxygen to make a protective shield of ozone. The ozone layer is below mesosphere and the mesopause. The ozone layer was built up by the photo dissociation of molecular oxygen O2 into two single oxygen atoms. One of the single atoms combines with molecular oxygen to make ozone O3. Ozone absorbs ultra violet radiation. The increase in temperature, the temperature inversion layer in our stratosphere is caused by the vibration and rotation of ozone molecules heated by the UV radiation from the Sun. In other words. it does not matter if we also had some cometary impacts. If we want a stable ozone layer and a lot of oxygen, the must be life and photosynthesis.
Could There Be Life in the Cloudtops of Venus?
AUGUST 21, 2020 BY NANCY ATKINSON
When it comes to places with the potential for habitability, Venus isn’t usually considered on that list. The hot, greenhouse-effect-gone-mad neighboring planet with a crushing surface pressure and sulfuric acid clouds certainly isn’t friendly to life as we know it, and the few spacecraft humanity has sent to Venus’ surface have only endured a few minutes.
But up about 40 to 60 km (25 to 37 miles) above the surface, the atmosphere of Venus is the most Earth-like of any other place in the Solar System. There, Venus has air pressure of approximately 1 bar and temperatures in the 0°C to 50°C range. It’s not quite a shirtsleeves environment, as humans would need air to breathe and protection from the sulfuric acid in the atmosphere. Plus, also consider that Venus is considered to be in the habitable zone of our star.
Full article here:
https://www.universetoday.com/147509/could-there-be-life-in-the-cloudtops-of-venus-2/