The seven planets circling the star TRAPPIST-1 have been lionized in the media, and understandably so, given that more than one have the potential for habitability. But of course M-dwarfs call up the inevitable problems associated with such tiny stars. Habitable planets must orbit close to the star, with the probability of tidal lock and subsequent climatic issues. Moreover, the flare activity particularly in young M-dwarfs gives cause for concern.
It’s the latter issue that Jack T. O’Malley-James and Lisa Kaltenegger (both at Cornell, where Kaltenegger is director of the Carl Sagan Institute) have explored in a new paper to be published in The Astrophysical Journal. As the paper explains, the question of habitability becomes troubling when we realize how frequently an M-dwarf can flare. Proxima Centauri, an M5 star, undergoes intense flares every 10 to 30 hours, with effects on the planet in its habitable zone that are still unknown. Can a planet with high doses of ultraviolet radiation remain habitable under such bombardment?
We probe such questions with particular urgency because 75 percent of the stars in the neighborhood of the Sun are M-dwarfs, and we’re about to send the TESS mission (Transiting Exoplanet Survey Satellite) into space to look for habitable zone planets around nearby stars. TESS will be able to do this for late M and early K stars, and as Kaltenegger notes, the mission is expected to uncover hundreds of planets between 1.25 and 2 Earth radii in size, and tens of Earth-sized planets, some of them likely to be in the habitable zone.

Image: Artist’s impression of the Alpha Centauri stellar system as viewed from the surface of the habitable-zone planet Proxima b. Credit: ESO/M. Kornmesser
TESS findings will be handed off to future ground and space-based missions for detailed study, making it likely that the first habitable zone planet that we can characterize will orbit a nearby M star. Recent studies have looked at the biological effects of radiation for atmospheres corresponding to different times in the evolution of the Earth, finding that such a planet orbiting an inactive M-dwarf would receive a lower ultraviolet flux than Earth.
But active M-dwarfs are another story. Planets orbiting such a star would receive bursts of UV in their tight, habitable zone orbits, increasing the surface UV flux by as much as two orders of magnitude for up to several hours. Add in the fact that M stars remain active for longer periods than G-class stars like the Sun, and the scenario looks dicey for life indeed. Consider the atmospheric effects, as noted in the paper:
The close proximity of planets in the HZ of cool stars can cause the planet’s magnetic field to be compressed by stellar magnetic pressure, reducing the planet’s ability to resist atmospheric erosion by the stellar wind. X-ray and EUV flare activity can occur up to 10-15 times per day, and typically 2-10 times, for M dwarfs (Cuntz & Guinan, 2016), which increases atmospheric erosion on close-in planets. This results in higher fluxes of UV radiation reaching the planet’s surface (Lammer et al. 2007; See et al., 2014) and, potentially, a less dense atmosphere.
We also have to keep in mind that in such close orbits, the effect of the star’s stellar wind would be orders of magnitude stronger in the habitable zone than what Earth experiences. The result: Erosion of any protective ozone shield and potential loss of at least some of the atmosphere. Biological molecules under such conditions can undergo mutation. We’re fortunate that on our planet, the ozone layer blocks the most damaging UV wavelengths. For life to flourish, some way of protecting it from this radiation is thus a prerequisite.
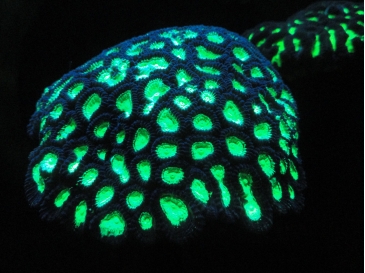
Going underwater is one solution, or burrowing deep into the ground, a strategy that could prove life-saving but also make detection by remote instruments extremely difficult. But there is one adaptation that would be detectable: photoprotective biofluorescence. In this process, protective proteins absorb harmful wavelengths and re-emit them at longer, safer wavelengths. Some species of coral are believed to use this mechanism to protect symbiotic algae needed for energy, with fluorescent proteins absorbing blue and UV photons and re-emitting them at longer wavelengths.
Image: This is Figure 1 from the paper, showing an example of coral fluorescence. Coral fluorescent proteins absorb near-UV and blue light and reemit it at longer wavelengths (see e.g. Mazel & Fuchs 2003). Image made available under Creative Commons CC0 1.0 Universal Public Domain Dedication.
Biofluorescence occurs when specialized fluorescent molecules are excited by high-energy light, causing them to absorb part of that energy and release the rest at lower-energy wavelengths. Such biofluorescent light can only be seen when the organism is being illuminated, which could be the key to their detection on other worlds. From the paper:
Photoprotective biofluorescence (the “up-shifting” of UV light to longer, safer wavelengths, via absorption by fluorescent proteins), a proposed UV protection mechanism of some coral species, would increase the detectability of biota, both in a spectrum, as well as in a color-color diagram. Such biofluorescence could be observable as a “temporal biosignature” for planets around stars with changing UV environments, like active flaring M stars, in both their spectra as well as their color.
The color-color diagram referred to above is frequently used in the study of star-forming regions, and as a way of comparing the apparent magnitudes of stars. Here one observes the magnitude of an object successively through two different filters — the difference in brightness between two bands at certain wavelengths is what is referred to as ‘color.’ One brightness range is plotted on the horizontal axis, while another range is plotted on the vertical axis.

Image (click to enlarge): A color-color diagram. This is Figure 10 from the paper. Caption: Color-color diagrams of planet models with an atmosphere and 50% cloud cover, and a surface that is completely covered by biofluorescent corals, fluorescent minerals, or vegetation compared to the colors of planets in our own Solar System, before (grey – labelled A, B, C, D) and during fluorescence at each of the four common emission wavelengths. Credit: Jack O’Malley-James / Lisa Kaltenegger.
The authors argue that color-color diagrams can distinguish biological sources of fluorescence from abiotic ones, as the change in color that biofluorescence produces differs from all other forms of the phenomenon:
Using a standard astronomy tool to characterize stellar objects, a color-color diagram, one can distinguish planets with and without biofluorescent biosignatures. The change in color caused by biofluorescence differs, in position and magnitude, from that caused by abiotic fluorescence, distinguishing both.
The authors point out that Proxima b will be a prime target to search for biofluorescence with the upcoming European Extremely Large Telescope. But TESS should give us a number of useful targets as we examine stars during flare periods looking for a biofluorescent biosphere. In other words, it may be that in the case of a planet orbiting an active M-dwarf, it’s precisely the high UV flux that will allow us to detect a biosphere that would otherwise be hidden.
The paper is O’Malley-James and Kaltenegger, “Biofluorescent Worlds: Biological fluorescence as a temporal biosignature for flare stars worlds,” to be published in The Astrophysical Journal (preprint). A second paper from Lisa Kaltenegger, “UV Surface Habitability of the TRAPPIST-1 System,” is now under review at Monthly Notices of the Royal Society. More on that one when I have a copy to work with.



This is innovative work no question about that. For those who are interested in the genesis of ideas though, it has to be noted that during a lecture on astrobiology at the University of Heidelberg in late 2013 I presented my concept of convergent evolution of bioluminescence on Europa (see my essay “The Blue Spectres of Abyssal Europa” published here on CD) based on the top-down selection pressure of oxygen defense (mirrored here by UV radiation) during one of the lectures held by Prof. Kaltenegger. I provided a copy of my abstract, detailing those concepts, for the “Habitability of Icy Worlds Workshop” in Pasadena:
http://www.hou.usra.edu/meetings/icyworlds2014/pdf/4040.pdf
The preprint makes reference to “convergent evolution arguments” multiple times without detailing what these arguments are actually saying. Convergent evolution arguments and their relation to astrobiology are found in my 2015 article “Convergent Evolution and the Search for Biosignatures within the Solar System and Beyond” where I give a comprehensive account on the published literature in the field (e.g. the fine work of Conway Morris or Chela-Flores). The bioluminescence idea is laid out in detail in this paper as well.
I got in touch with the authors pointing out this “deep homology” (in contrast to bona fide convergent evolution of ideas, since Prof. Kaltenegger should, in any case, have remembered my presentation of bioluminescence when speculating on biofluorescence). Let’s see if the reference to bioluminescence, as promised, will find its way into the final version of the paper.
The “glowing aliens” of Proxima Centauri B and beyond have created quite a stirr in the media, including a news item in one of the biggest German magazines. Well, I was the first one to specifically publish and present on the possibility of (convergent) glowing aliens, as can easily be proven:
http://sciencev2.orf.at/stories/1732985/index.html
“Galactic Fire Flies”
Frankly, I am finding it quite unprofessional for an experienced researcher to take credit (especially in the media) for brilliant, original ideas (or rather variations of already existing concepts) that have been already advanced by others without properly referencing the existing state of the art.
I would not make the present case had I not presented my idea during a lecture of Prof. Kaltenegger detailing the process of constraining biosignatures based on convergent evolution. However, I’d like to hear more about the genesis of their idea from their perspective. But a statement as the following is not acceptable, as others have demonstrably, with the author’s knowledge, thought along these lines and even published on such ideas.
“We decided to look into more advanced organisms and see if they had any mechanisms that could produce signs of life nobody thought of yet,”
http://news.nationalgeographic.com/2016/08/planet-proxima-centauri-life-glowing-corals-radiation-space-science/
I am prepared to stand my ground on this matter, but the issue had to be clarified.
Best regards,
Claudio L. Flores Martinez
Life is highly resilient and adaptable. I believe that an alien biology could evolve to cope with flares/radiation, just as life on earth has evolved to cope with many different challenges. My main concern is the effect on the atmosphere — if flaring could strip away atmospheres, that could destroy habitability. Other than that, the direct impact on life is a non-issue imo.
The concept of biofluorescence is somewhat ironic given the old direct imaging metaphor of “looking for a firefly by a search light “. Quite literally now too.
My concern about this is that although I can see such a biological adaptation arising from natural selection in the same way as photosynthesis, and how it might also analogously be incorporated into multi cellular life ( coral analogues et al ) -as with cyano bacteria into eukaryotes – but what would the starting point be on an M dwarf planet stripped of its atmosphere by a hostile and relabelled high energy stellar flux ? Evolution needs something to work on , as opposed to full biofluorescence arising spontaneously .
Another issue is that it is a long way from absorbing reactively low energy UVA and fluorescing longer wavelengths on Earth after long periods of evolution out of the extreme fringes of a robust biosphere , to absorbing much more energetic EUV overlapping into the X-Ray band from a tenous and challenged planetary ecosystem trying to gain a foothold . Likely clinging onto existence under ground or at best deep under water with both hardly accessible to UV and especially the less energetic version that might kick start the process .
Extremophiles on Earth haven’t just arisen in immediate response to a hostile environment but are the hardy survivors of a generic biota benefiting from years of gradually increasing exposure induced evolution which allows slow expansion into that environment.
As long as life could get started and attain a foothold underground or underwater, it would be fine. It could slowly migrate closer to the surface, with less rock/water shielding, and gradually adapt to increasing levels of radiation — eventually living freely on the surface. The key to this is a subterranean environment to act as an incubator for the biosphere.
Xonobiology https://en.m.wikipedia.org/wiki/Xenobiology
could prove to be weirder than we imagine. Xeno nucleic acids https://en.m.wikipedia.org/wiki/Xeno_nucleic_acid and alien amino acids would not be so constrained http://www.space.com/24460-alien-life-building-blocks-amino-acids.html by Earth biology’s Last Universal common Ancestor, LUCA.
Bioluminescence and biofluorescence may well be ongoing phenomona “out there”, but their trains may or may not ride our tracks.
I agree with Ashley Baldwin. Life has to be able to survive past the small size or single celled and multi cellular size before it can evolve into a higher form and develop an adaptation to extreme environments like extremophiles and fluorescence which might be difficult in an environment with high UV and x-ray radiation.
I somewhat agree with Ashley. UV tends to break up organic molecules making it possibly very difficult for life to start assuming the “warm shallow pool” model. However, this would not be the case if life is formed in the deep ocean vents and protected from UV.
What I think is more likely that any life will not emerge into the atmosphere. If bioluminescence did evolve as a protective device, it would perhaps have helped organisms to reach nutrients in the UV exposed, upper regions of the oceans. Whether it could ever evolve to a capability to resist a lot of hard UV is a question. I am skeptical that any organic molecule can absorb the UV without breaking apart rather than just emitting it at a different wavelength. Perhaps a chemist could answer that.
Very good article, on worlds that are tidally locked you should still a
have librations that could create much better habitability zones near the terminator. For example TRAPPIST-1 planets would have large tides and librations because of the planets passing so close to each other. The UV and X-rays would be have been filtered by going thru 100’s of miles of atmosphere and ocean waters near the terminator. How about shellfish and clams, plus some of them burrow into the sand, that would protect from UV.
The rest of life on other worlds may find the earths oxygen an extreme environment and that we are extremophiles. Its all a matter of perspective and right now we have a very limited understanding of what these worlds are like. You can see the diversity that life has and has had on the earth to realize that there are more forms of life out there then we can imagine.
The new K2 data has been released for TRAPPIST-1:
https://www.jpl.nasa.gov/news/news.php?release=2017-060&rn=news.xml&rst=6767
http://archive.stsci.edu/k2/trappist1/
Raw data, anyone know a good article on proccesing this?
Some interesting articles on AI and image processing for exoplanets:
Generative Adversarial Networks recover features in
astrophysical images of galaxies beyond the deconvolution
limit:
https://www.wired.com/2017/03/astronomers-deploy-ai-unravel-mysteries-universe/
https://arxiv.org/pdf/1702.00403.pdf
Meet RobERt, the dreaming detective for exoplanet atmospheres:
https://www.sciencedaily.com/releases/2016/06/160627214628.htm
https://arxiv.org/pdf/1511.08339.pdf
I agree with Ashley Baldwin, Geoffrey Hillend and Alex Tolley about the problems with life *arising* under a huge flux of UV, and that it would probably have to arise in more sheltered conditions first.
However, lets speculate..
1.) Maybe lifeforms could actively USE the UV for energy. This doesn’t require some fiercely UV-resistant organic molecule, a UV-hardy flourescent mineral would suffice (the organism could accumulate it around itself if it already had some energy), and feed photosynthetic processes (sort of what the article alludes to),
2.) To venture out into the high UV, couldn’t an organism secrete an *inorganic* shell (say silicate or carbonate as happens on Earth) rather than a UV-sensitive organic covering? It would likely have to secrete a shell while still in a sheltered place (or inside a parent or egg?).
For extra points, combine (1) and (2), a UV-resistant ‘shellfish’ with a flourescent shell, and photosynthetic organs within, for instance.
I guess such an inorganic flourescent structure would be hard or impossible to distinguish from ordinary mineral deposits, though. Also, an ocean’s (?) chemistry would have to allow shell formation (now a bit of a worry on THIS planet). Also, would it be difficult to have small planktonic organisms, as their shells would have to be heavy relative to their size ?.
Michael T
So what I think you are saying is any even a photosynthetic organism would need a surface shell to protect even the bioluminescent protein layer that converts the energetic sunlight to wavelengths suitable for chlorophyll-like macromolecules to process.
As long as the organisms are aquatic, gases in the shell could keep the organism buoyant and even adjust the depth if the radiation gets too hard.
On land, this would be a problem, unless the “plants” could move to shelter.
If life evolved in such a world, it would probably have to do so in very sheltered conditions, either in the deep ocean or possibly underground in some way. I cannot see how it could evolve if exposed to such hard radiation, as the organic molecules would keep breaking apart.
If life evolved, remained unicellular and lived in the deep ocean, it could still generate gases detectable in the atmosphere. Such a world could have had its “Great Oxygenation Event” so that we could detect an abundant O2 atmosphere. However, could that be distinguishable from the O2 generated by greater photolysis?
@ Alex Tolley,
Thanks for your reply.
No, I was thinking of inorganic fluorescent materials (phosphors?) in an inorganic shell, and then organic photosynthetic layers underneath the shell, using the secondary light (+ sunlight). I don’t see why the material that does the fluorescence has to be organic; there are inorganic candidates. I know the paper was about organic ones.
This is assuming an organism uses photosynthesis as well as just using fluorescence for wavelength conversion. The reason I discussed photosynthesis is that the corals that use fluorescence have photosynthetic symbionts. An organism might have symbionts or do its own photosynthesis. I do rather like my photosynthetic shellfish, though :-)
Aye re bouyancy imparted by gases in a shell, didn’t think of that. Or use some thing akin to a fish’s swim bladder or the depth control mechanism used by Nautilus, etc.
I agree re evolving in shelter first. As for shell development, it just takes some lucky mutation leading to a bit of shell accumulation, giving an organism just that little bit of advantage in UV, and a little bit more range. Something similar must have happened for shelled creatures on Earth.
I agree re oxygenation. I didn’t address it because I was still thinking of direct biofluorescence detection.
[ Dashed typos! It’s “fluor”, not “flour”, unless the aliens make bread out of it :-) ]
Mike
An interesting twist to the “Rare Earth” hypothesis and as far as I recall, not something they considered when the book was published, UV flares and atmosphere/ozone layer depletion.
Are we gathering evidence to support their conjecture… that micro-biota may indeed be rampant throughout the cosmos…. but there are special conditions that require multi-cellular, “higher” forms of life?
UV might not be a problem, but the hard x-ray outbursts constantly coming off a Type-M are going to cook any macromolecules into cross-linked muck. Along with atmospheric speckling from the outburst’s accompanying proton flux, it’s more than likely you’re looking at a half dozen Mercury and Venus look-alikes. If you’re a layperson, Ward & Brownlee’s book, “Rare Earth” is worth a read, as is the Chandra investigation group’s explanation of why M-Type stars are horrible hosts for planets. http://chandra.harvard.edu/chronicle/0407/RedDwarf/
Scott the Orion star cluster is very young, if you check the original data they say 1 to 10 million years. These red dwarfs are very active flare stars because of the very young age. The orion nebula is one the best nearby star forming regions and an excellent target for small telescopes and 7×50 binoculars.
Yes, X-rays bring on even more problems than UV. I can imagine shells shielding against UV but not X-rays or other radiation.
Organisms would have to be in caves, crevices or deeeeep in oceans.
And I agree with the other commenters above with regard to all the problems concerning habitability and (higher) life arising on planets around M dwarfs, also recalling the recent post A New Look at Habitability around Red Dwarf Stars, about ‘space weather’, a combination of X, UV, stellar wind, and the like.
https://centauri-dreams.org/?p=37109#comments
http://iopscience.iop.org/article/10.3847/2041-8213/836/1/L3
Quote: “the authors estimate oxygen loss would occur in its (Proxima b) atmosphere within 10 million years”.
As I said before, I think it is no coincidence that we orbit a G star, despite this spectral class being relatively uncommon. Statistics based on 1 are totally unreliable, but it may be telling.
Tardigrades handle UV quite well, short to medium bursts look quite liveable to these animals.
http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0064793