Centauri Dreams regular Alex Tolley here examines a new paper with a novel take on Saturn’s moon Enceladus. Tempting us with its geysers and the organic compounds Cassini detected in their spray, Enceladus offers the prospect of life within its internal ocean. But are there other explanations for what we see, pointing to what may be a prebiotic environment? For that matter, what features of life’s chemistry could emerge on such a world without yet maturing into what we would recognize as living organisms? The paper Alex examines offers us quite an interesting take on a possible origin for life not just on Enceladus but elsewhere in the universe.
by Alex Tolley

Image: “Snow on Enceladus.” Credit: David Hardy.
The discovery of subsurface oceans in the icy moons of Europa and Enceladus has increased interest in the exploration of these moons. The logic of the mantra “Follow the water” implies that there may be extant life in these oceans, most excitingly from a unique genesis at hypothesized ocean hot vents that release the tidal heat. The Europa Clipper is one such Probe.
A new review article published in Astrobiology by Amit Kahana (Weizmann Institute of Science, Israel) and colleagues makes a case that rather than being biotic, Enceladus is prebiotic, a state such as has been speculated on Titan. As the authors state:
The enceladan subglacial ocean appears to be the first observationally discovered concrete example of what may well be a primeval soup.
They come to this conclusion via a chain of logic which follows.
The first piece of evidence is that the Cassini probe detected organic compounds in the plumes of Enceladus and Saturn’s rings. This should not be surprising as we might expect organic compounds as they are common on icy bodies like comets, which include the primordial compounds that can be converted to compounds that are presumed to be the basic building blocks of life on Earth. They tabulate 53 compounds identified in space, most of which are organic with up to 10 carbon atoms. There is also abundant insoluble organic matter (IOM), like kerogens that can be broken down into small molecules. The authors interpret the Cassini data to suggest the detected organic compounds may have up to 150 carbon atoms, similar in composition to the IOM detected elsewhere. The authors believe that the these organics are rich in carbon and hydrogen, and depleted in oxygen and nitrogen. This indicates methane and other alkanes, longer chain organics or aromatics, rather than the mix of organics that might be expected if life was extant..
On the other hand, the observations are consistent with a predominance of relatively long unsaturated hydrocarbon chains, aliphatic or aromatic, with a small number of polar heteroatoms.
The authors take pains to suggest that the origin of life research is dependent on the primordial compound mix and the energy sources to transform them to prebiotic compounds. For example, they counter the requirements of the classic Miller-Urey experiment that added electrical energy to reducing gas mixtures to create amino acids. They contrast the results of this experiment with the speculated abundance and composition of organics in the plumes of Enceladus to suggest other means of production. These organics could have been delivered by comets, or created ?de novo? within Enceladus, beneath a thick ice crust, a very different environment from that on the early Earth. The authors paint a picture that favors Oparin’s primordial soup model in which heterotrophic early life feeds off the energy in the organic molecules in this soup. [Heterotrophic life consumes these “soup” compounds, rather than autotrophic life, such as methanogens which consumes inorganic CO2 and hydrogen to extract the energy from methane (CH4) synthesis, as we see in many Archaea extremophiles.]
A primordial soup is the model they have of Enceladus, with organics delivered by infall, with perhaps a rich layer of IOM between the ocean surface and the ice crust above (see figure 1 below).
If the ocean contains mainly high molecular weight organic material, then how are the lower molecular weight organics created? Experimental work suggests that it is the energy, and possibly mineral catalysts, at the hot vents that convert the IOM to the smaller molecules:
In parallel, high-temperature hydrous pyrolysis experiments of solid fossil fuels such as kerogen and bitumen, in the presence or absence of mineral catalysts, gave rise to a plethora of low-molecular-weight organics. The products included alkanes, alkenes, and isoprenoids (Huizinga et al., 1987) as well as long-chain fatty acids (Kawamura et al., 1986; Siskin and Katritzky, 1991). In another set of studies, hydrous pyrolytic degradation of petroleum sediments (kerogen) has been examined both in hydrothermal vents and in the laboratory (Leif and Simoneit, 1995). A major set of products constituted amphiphilic n-alkanones with chain lengths from C11 to C33. A similar set of results showed how hydrothermal pyrolysis leads to the formation of diverse polar compounds, including alkanoic acids and alcohols, isoprenoid ketones, and alkanoate esters, in the C9–C33 range (Rushdi and Simoneit, 2011). All these results are consistent with the possible generation of monomeric organics from insoluble polymers under conditions similar to those prevailing on Enceladus. Whether this actually happens on Enceladus could only be verified by future missions and analyses.
The authors’ model is that of an ocean rich in organics primarily from impactors. Much of this material is IOM. The silicate material detected in the plumes infers that there are hot vents on the ocean bottom that help break down the IOM into simpler, C, H rich molecules, which can then form the lipid membranes of micelles and vesicles. It is the mix of vent emissions and these lipids and IOM that Cassini detected in the plumes. The apparent paucity of nitrogen and oxygen in the plume organics leads the authors to suggest that the compounds likely have an abiotic origin, although they do not rule it out:
However, in the absence of evidence to the contrary, a biotic origin of Enceladus’ organics remains a remote possibility (Postberg et al., 2018).
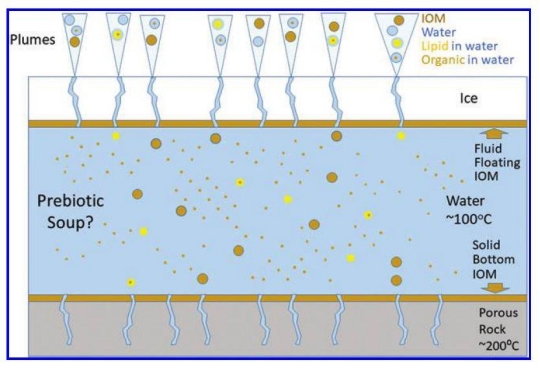
Figure 1. Enceladus cross-section, showing the different potential components of the soup and the plumes. IOM stands for insoluble organic matter, which is taken to be synonymous with nondispersive organic matter. Such polymeric compounds can still be carried in the plume as small particles detached from insoluble organic layers at the top of the water layer. Lipid in water alludes to monomers and aggregates such as micelles and vesicles. Organic in water is dispersed, for example as a microemulsion. Inspired by the data in Postberg et al. (2018).
Figure 1 above shows the authors’ model of the organic chemistry of Enceladus.
Given the authors’ assumption that abundant lipids exist, and that they have a mechanism for creating them, what does this mean for abiogenesis and life? The two most dominant theories for the origin of life are the? RNA First? and Metabolism First? theories. Both can exist without surrounding cell walls to contain either the RNA (as both catalysts and information molecules) or the metabolites for autocatalytic sets. However, some form of concentration and protection from degradation is eventually needed, and terrestrial life used lipid membranes to provide that solution. However, there are other theories, and Lipid First? is more akin to the idea that lipids can form mono and bilayer membranes that can then encapsulate the metabolisms and replication machinery of the protocells. In such a world, we might expect to see vesicles or coacervates that concentrate the needed components for the transition to life.
The authors argue that the best origin of life model is one that fits with the observed chemical environment, rather than with a predetermined set of molecules and energy sources such as the Miller-Urey experiment required. They use their speculated set of lipid precursors to define the organic soups and from this favor the Lipid World hypothesis. In other words, having built up a case for long-chain unsaturated carbon molecules from their interpretation of the data, they use this as the given compound environment to base their origin of life scenario. They then argue that cell reproduction occurs by membrane splitting and that information about the membranes was added by the later incorporation of informational molecules like RNA. Thus their Lipid World would predate the RNA world and Metabolism World in this scenario.
Assuming that the authors are correct and Enceladus’s ocean contains a “Lipid World”, the next question they ask is this even prebiotic? It is generally accepted that lipids can form abiotically and organize into vesicles or micelles. Can the lipid membranes help catalyze reactions? A paper very recently published [1] suggests that amino acids can bind to lipid membranes, both stabilizing them and providing the means to catalyze reactions such as peptide formation. This would then allow the concentration of metabolites into these protocells. The experimental results from this paper would support a Lipid First model of the origin of life.
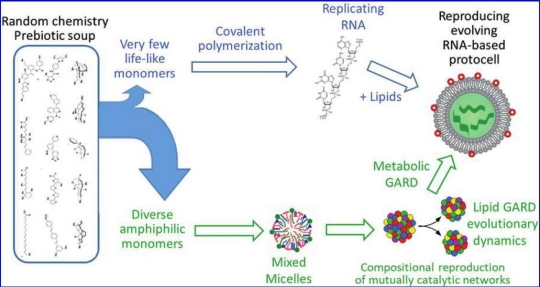
Figure 2. From soup to protocells, delineating the underpinnings of two different models, RNA First (RF, top) and Lipid First (LF, bottom). In RF, specific monomers are singled out from a heterogeneous chemical mixture and undergo polymerization, culminating in the emergence of a self-replicating polymer. This is then enclosed in a lipid bilayer, leading to protocells capable of selection and evolution. In LF, a large variety of amphiphiles spontaneously generates a plethora of assemblies, for example, micelles. The GARD model then predicts that very specific micellar compositions establish a mutually catalytic network, which may exhibit homeostatic growth. This, when followed by fission events, constitutes a reproduction system, capable of selection and evolution (see ‘‘How do lipids reproduce?’’ Section 8.4 and Lancet et al. [2018]). Subsequent prolonged evolution may lead to the emergence of RNA, proteins, and metabolism (See Section 8.5, ‘‘How would lipids beget full-fledged protocells’’ and Lancet et al. [2018] Section 11.1). Caption source: Kahana et al.
Figure 2 above shows the authors’ view of the Enceladus Lipid World hypothesis that would culminate in prebiotic protocells, but not fully living cells.
If their hypothesis is correct, then Enceladus is in a prebiotic state. The components to start life may be present, but mostly concentrated in protocells, but not in any form that we would call life. They may show some features of life, such as motility and replication by division, but no true metabolism to liberate energy for growth and reproduction, nor the information molecules needed to encode the instructions that can evolve under natural selection to drive these protocells to transition to living cells. Such a state would still be a fascinating one for study as it would offer a prebiotic environment that has long been lost to us once life emerged on Earth.
However, it should be borne in mind that their argument is based on a chain of logic. Any link that proved false could break that chain. The organic material may not be as depleted in nitrogen and oxygen as inferred. The organic material in the plumes may not be as complex as believed. The ocean may not have a lot of IOM as a feedstock, and reactions may not result in lipid chains, but rather a mix of different molecular weight organics, both linear and aromatic. The organics may therefore be primordial rather than prebiotic. The Cassini data are tantalizing and beg for further missions to elucidate the nature of this moon. Hopefully we may get further clues from the Europa Clipper mission.
The paper is Kahana A et all “Enceladus: First Observed Primordial Soup Could Arbitrate Origin-of-Life Debate”, Astrobiology v12 (10) 2019. Full Text.
References
1. Caitlin E. Cornell, Roy A. Black, Mengjun Xue, Helen E. Litz, Andrew Ramsay, Moshe Gordon, Alexander Mileant, Zachary R. Cohen, James A. Williams, Kelly K. Lee, Gary P. Drobny, and Sarah L. Keller. ? Prebiotic amino acids bind to and stabilize prebiotic fatty acid membranes?. ? PNAS 116 (35), (27 August, 2019), 17239-17244 (abstract).



Speaking of strange substances on a moon…
https://www.space.com/china-far-side-moon-rover-strange-substance.html
Whose idea was it to use a black and white photo of the “strangely colored substance”?
As far as I know they showed the crater the substance is in, but no images of the actual “gel” yet.
Any updates from those in the know would be appreciated here, thank you.
The general topic is fascinating. Thank you Alex. I am sadly incapable of forming an intelligent opinion on it for lack anything above very basic knowledge of organic chemistry.
It is however notable that Enceladus will begin transiting Saturn again in 2022. That offers the potential to deepen our data set on the plume composition. Assuming that the noise (from Saturn) can be filtered out.
Regarding the question of whether life may exist there, I do wonder at the timescales required under the local conditions for the emergence of life or even a protocell complete with some (waves hands) informational polymer . If the moon is indeed as young as some have posited, there may have simply not been sufficient time for life to emerge. 30, 60 or 100 million years. Enough time?
Freely available:
Biochemistry
An Introduction to Molecular Biology
With regard to the moon, the absence of the necessary substrates and milieu would preclude abiogenesis until the heat death of the universe. It does not preclude the emergence of intelligence from “unknown unknowns” (unknown unknown to our knowledge and/or scope of awareness).
We really don’t know how long it took for life to appear on Earth. It seems as though we find microfossils fairly soon after Earth has acquired its liquid oceans and the bombardment has stopped. That implies that life appeared fairly quickly. Of course, we cannot rule out some sort of panspermia, although that just opens up more questions on origins.
Rather than speculate, I would prefer we go and look. We should look both elsewhere in our solar system and the rich haul of exoplanets. In our solar system we have the benefit of being able to analyze samples, but the problem of a few targets. Exoplanets limit our analytical techniques and the information we can get, but there are potentially many more targets and some that may prove Earth-like.
I’m not sure if you’re suggesting phospholipids or lipids account for the 800-1250 peak – the idea is intriguing. However, as speculations go, there is something I prefer over the notion of empty lipid membranes floating in solution.
According to Usami and Okamoto ( https://pubs.rsc.org/en/content/articlelanding/2017/OB/C7OB02051A ) the formose reaction (converting five formaldehyde molecules into a ribose) can be catalyzed by hydroxylapatite, the mineral of bone and tooth. Our bodies have an affinity for hydroxylapatite, making it even in places like the coronary arteries where they shouldn’t; even our old cesspits turn into carboxylated hydroxylapatite. This is because the cell membrane phospholipids can fit up against its crystal surface.
So I’d imagine a prebiotic lawn of ribose, glyceraldehyde and other small CH2O compounds forming on hydroxylapatite, which -somehow- recruit nitrogenous bases to make potential nucleotide cofactors including some with a folate-like ability to store formaldehyde. Krebs cycle intermediates (dicarboxylic acids) chelate the calcium ions. Loose carbonate and divalent cations (Fe, Mg, Zn) might be present. The system develops a means akin to the Krebs cycle to catalytically rearrange carbohydrates to release CO2 while leaving something closer to fatty acid bound to the phosphates.
At first this fatty patch might merely adsorb nutrients, but once some of the underlying mineral dissolves, it leaves a tent of cell membrane overlying a rich cellular metabolism. The system can enjoy some benefits of isolation without locking itself inside a membrane completely. The first nucleic acids might assist the original biochemistry by genetic assimilation, and large lipid deposits might fold in on themselves to release free ‘cells’ to spread the colony to remote mineral sites.
Mineral surfaces to help form and concentrate materials is an attractive idea. that has been explored, albeit mostly theoretically. The paper you cite would suggest that hydroxyapatite would act as a catalyst to produce ribose that could then form the backbones for RNA (RNA World).
My thought is that if this reaction is possible on Enceladus (or Europa), then we might expect to see ribose in the plumes. We really need a good analysis of the plume chemistry to get some handle on the subsurface ocean chemistry.
I don’t know the field well enough to know what to ask for. Looking at the Kahana paper I am tempted to claim the first three peaks of “90-200 u” are glyceraldehyde, dihydroxyacetone, and ribose … an exercise in hallucination, to be sure! But to take a step back, I don’t even know that hydroxylapatite is on the menu for Enceladus. If the mineral is at the bottom of the ocean (which I don’t know), it should be able to tolerate the heat and the pressure, but as little as 100 C might make ribose accumulation impossible ( https://www.pnas.org/content/92/18/8158 ). Do we know how deep the rocky surface of the moon is, or how hot it is? (It seems like lot to deduce from one small probe in orbit)
I think one can resolve your issue here:
1. If hydroxyapatite exists, the plumes indicate that particles of rock are present. To me, this means that particles will be present dispersed in the ocean
2. Based on [1], this implies that there are catalytic surfaces for ribose to form in a variety of temperatures and that the temperature range of the ocean should provide for the accumulation of these molecules.
This is just my guess, as I am not a chemist.
The model from the image above proposes an ocean of 100 C water under a thick layer of ice, which at 0.11 g it should be ordinary ice Ih. A layer of insoluble organic matter that is presumably lighter than water provides insulation, but multiple geysers pass through it and tectonic resurfacing is occurring. If the IOM is volatile, it seems like it should flow out easily, leaving hot water and ice to work out some arrangement. If the IOM is not volatile, it should come up in chunks as the ice above it is melted away, spraying the surface of the moon with dark pebbles and exposing a fresh ice surface to the boiling water below. Either way, I can’t see how this could be stable.
I think the authors have either misinterpreted the Postberg (2018) paper or done a poor job with the graphic in Figure 6 (of their paper) and Figure 1 in my post. This is what Postberg had to say:
IOW, the 100C is at the ocean floor boundary, and that mixing allows all the ocean water to be processed at 100C, not that the ocean is at 100C. I would be astounded that the ocean temperature could be maintained at 100C where it is in contact with the ice crust, whatever insulation it may have. It would also imply 100C water being pushed up through the cracks in the crusts which would surely erode those cracks very quickly.
Thanks for tracking that down! This scenario seems more plausible and hopeful for life: earthlike ocean water, earthlike ocean pressure and temperature, with geothermal vents at the bottom. We practically know life can survive under such circumstances, and some argue it started there. The solutes may be different, and I don’t know how low the gravity can be before black smokers start autoclaving everything within dozens of meters instead of forming nice chimneys you could approach in a submarine. Still, diffuse geothermal seeps are nearly as interesting, and I don’t see how low gravity could interfere with those.
Thanks for another insightful piece, Alex!
Whether RNA First, Metabolism First or Lipid First, sooner or later they all had to come together so that I can comment here.
External influences could also have played a significant role.
Did clay mould life’s origins?
Clay may have been birthplace of life on Earth, new study suggests
Great article Alex,
Understanding there are many competing needs for science dollars, it seems to me that orbital surveys, landers, and return missions to the icy moons Europa, Callisto, and Ganymede, should be prioritized.
Would not the findings would deeply inform our understanding of JWST and other large telescope data as they begin to survey exoplanet surfaces and atmospheres for signs of life?
Just wish it would happen in my lifetime!!
I’d be happy to advocate the stance that if Enceladus is an example of a prebiotic environment that is potentially partway along the road to life, this is a better scientific opportunity (though not so tabloid headline friendly) than an actual extraterrestrial life form with billions of years of evolution behind it. The rationale for seeking out alien life is, after all, to better understand the origins of life generally, and hence ourselves. To some extent Enceladus could be representative of early solar system worlds, such as Ceres or other asteroids and dwarf planets with a history of subsurface water and organic chemistry interacting in the deep past, when they were warmer
Finding a prebiotic environment would certainly be a prize. However, we might still be presented with a problem – does it represent our prebiotic state or just one of many possibilities? OTOH, finding different evolved ET life would help to determine what is the space of biology, biochemistry, and molecular biology. It really depends on your interest area.
It may just me, but it seems intuitive that a prebiotic or probiotic system, studied in depth, could better demonstrate the possible space into which a water/carbon biochemistry could evolve than an evolved organism or biosphere – one can still demonstrate the fundamental biochemical possibilities arising from the initial mix, the other will have had many at of them pruned by natural selection. We have seen hints of this in the evolved organic matter found in carbanacious chondrite meteorites, which show that a great many more amino acid and nucleobase combinations that life currently uses were present and interacting in early solar system environments – many of the specifics of which we’ve lost in today’s terrestrial biosphere, or any evolved biosphere we may find extant. If we have a great many initial prebiotic states to start from this is still true. What the prebiotic environment won’t give us is the specifics of what evolutionary solutions to various challenges might evolve… but that would indeed be most relevant to a very specific interest. A bona fide proto biotic environment would give us the context that we logically need first. And, of course, it won’t provide that PR friendly ‘we found aliens’ moment.
If I understand you correctly, you are saying that the prebiotic chemistry landscape is much broader than the one selected for by terrestrial life, e.g. possible amino acids. This may be correct, but it may also be limited by the specific conditions of our solar system and even the planet. What if there are many routes and choices to life, some very exotic? Even terrestrial life is highly path-dependent by chance. We would want to find different examples of prebiotic environments to check if they are indeed similar rather than diverse. But then we have the problem of determining if any of these prebiotic states will become biotic, rather than remaining in a prebiotic state.
OTOH, if we can find many unique abiogenesis examples, even just in a prokaryotic state, it tells us a lot about the constraints that may or may not result in extant life. Because the size of possible protein space is so vast, I would also expect life to have taken many different paths given the pressures of convergent evolution. But of course, we are still blocked from understanding the prebiotic states and abiotic conditions that led to these life forms.
Both studies are going to prove very interesting, and both require that we can get samples of prebiotic and biotic environments.
In practice, that means we won’t get there until we have some way of sending out interstellar probes to collect the needed samples and living with the time to get results. The limitations imposed by c are really frustrating. Which is why we need ET life to come to us. If panspermia occurs, then we need ways to capture these organisms, whether they are hitching a ride on dust grains or in larger bodies like ‘Oumuamua.
We have a great many potential prebiotic environments to study, from the early days of our solar system: At one point a great, great many small worlds, such as Ceres, or the parent bodies of carbonaceous chondrite meteorites, had conditions that fulfilled the basic requirements. These are fossilised and preserved, albeit in damaged state, through out the solar system. Discounting the possibility of something truly exotic on, say, Titan (I think that is such an exotic possibility that we’re into a different discussion there) what we lack to really understand them is an extant prebiotic environment we can study – a ‘live’ specimen if you will, to bring all the others to life. Relevant comparisons to existing systems is a vital part of understanding records of past ones. If Enceladus is indeed a prebiotic world then, boom, there we have it: We can start to make better sense of all the others, even those that don’t exactly match. That stays true even if Enceladus is heading down a route that wouldn’t lead to life. The scientific value is potentially enormous, all from one relatively accessible world – it’s low hanging and very juicy fruit, relatively.
Not that it has to be either/or – but of we are indeed following the scientific rationales laid out for the search for extant alien life it seems very compelling to me that extant protobiotic / prebiotic environments deserve to be up there with as much priority. And if we’ve actually found one that’s relatively easy to access, well a bird in the hand…
Someone just made an argument for Mars as a prebiotic world and that it was worth looking for evidence there too.
If I were in a mischievous mood I might make the argument that the various organic ices thought to be beneath the lunar poles constitute a prebiotic environment. :)
“The rationale for seeking out alien life is, after all, to better understand the origins of life generally, and hence ourselves.”
Who decided that understanding evolution is our chief reason to know about life in the universe? I think it’s just the chief reason people use to get funding for their projects because it’s safe to say.
I genuinely appreciate that suspicion: I suspect that ‘because it’d be so freaking cool’ is actually closer to the motivations of many promoters of the search for extant alien life. That is, in a roundabout way, my point.
Ahem, probiotic = protobiotic. Spellcheck blues. I think I could actually just have said: Natural selection kills off a great many possible evolutionary paths, many of which might have been valid and selective under different circumstances, and the triumphant path usually wipes out the remains of the failed ones leaving little evidence. So we really will need prebiotic /protobiotic systems that are independent from Earth to understand how life arises, and what the biochemical space it can evolve into is, at least as much as we need to find well evolved extraterrestrial organisms.
Misread the title as probiotic… a moon made of frozen yoghurt?
Yes, that would be a rather startling spin on things!
If as some have hypothesized, life on Earth got started when a visiting spaceship flushed its toilets into the primordial ocean, that too would be a probiotic: our Last Universal Common Ancestor may have been from out of this world, from “a place where the sun don’t shine”.
Thomas Gold’s “Cosmic Garbage” theory was proposed when we knew far less biology than we do now. If we were to propose it today, with LUCA being in their excreted microbiome, it would mean that the astronaut’s biology might be more primitive than ours. This I find unlikely. While we cannot root our tree of life, it does seem more likely that the methanogens appeared first on our Earth, rather than later and retreating back into anaerobic niches.
If panspermia has happened (or continues to do so), I doubt it is the equivalent of the Apollo astronauts leaving their excreta on the Moon.
OTOH, I find your comment is very funny. Perfect for snorting my morning coffee!
Forty years ago at the start of this month, Pioneer 11 became the first spacecraft to flyby the planet Saturn:
https://www.americaspace.com/2019/09/01/visiting-saturn-40-years-since-pioneer-11s-mission-to-giant-ringed-planet/
Alex,
Is it reasonable to suppose that a second genesis of life on Enceladus would use RNA/DNA, or, might it be more likely that another coding molecule arises instead? Are RNA/DNA the best coding molecules or are we just biased to think so because these polymers are used in terrestrial biology? Is the RNA/DNA combo just one of potentially many versions of coding molecules for life that may be available in the universe (e.g. it just happened to be the circumstances on Earth that led to their selection here)?
That is question of great interest. There are also variations of these molecules too before we get to more exotic possibilities. For example, there is a hypothesis that the first information molecules (DNA?) used just 2 bases, rather than 4, limiting the number of amino acids in peptides assuming a 3-codon code. This is one reason why finding true de novo geneses could help answer that question.
Any small ring of water molecules in hexagonal ice has a series of hydrogen bonds where O-H … O could be reversed to store a bit of information. Even within the narrow universe of DNA-like molecules, several DNA-compatible codes like PNA have been proposed, and the code can also be extended to six base pairs ( https://www.nature.com/articles/nature24659 ).
If RNA (which evolved first) has a special role, it should be in terms of evolvability. The four bases and parts of flavin and pterin and folate all have a three-carbon unit linked with a lone carbon bound to at least two nitrogens. (It is tempting to compare this with the three-carbon scaffold of phospholipids) If there were some biochemical context where all RNA bases would form spontaneously, we could envision the system emerging on other worlds.
The information storage and replication by DNA use the phenomenon of a double-strand to ensure copying fidelity as well as simple damage control. It is possible that RNA was the first information storage molecule that allowed translation into proteins, but this role was taken over by DNA. RNA tends to fold back on itself and bind to complementary sequences in its own sequence which allows it to form 3-D structures that can act like proteins. That it serves multiple roles in teh cell is one argument for an RNA-first world. Alternative information storage molecules would still have to work with other molecules that can copy and translate that code.
The curious case of the transgressing tardigrades (part 2)
In the second part of their examination about the recent controversy about undisclosed microscopic life included on a lunar lander mission, a group of experts examines the regulatory issues in the United States that come into play in this incident.
Tuesday, September 3, 2019
http://thespacereview.com/article/3786/1