Until we learn whether or not life exists on other planets, we extrapolate on the basis of our single living world. Just how long it took life to develop is a vital question, with implications that extend to other planetary systems. In today’s essay, Alex Tolley brings his formidable background in the biological sciences to bear on the matter of Earth’s first living things, which may well have emerged far earlier than was once thought. In particular, what was the last universal common ancestor — LUCA — from which bacteria, archaea, and eukarya subsequently diverged? Without the evidence future landers and space telescopes will give us, we remain ignorant of so fundamental a question as whether life itself — not to mention intelligence — is a rarity in the cosmos. But we’re piecing together a framework that reveals Earth’s surprising ability to spring into early life.
by Alex Tolley
Once upon a time, the history of life on Earth seemed so much simpler. Darwin had shown how natural selection of traits could create new species given enough time, although he did not argue for the origin of life, other than it would start in a “warm pond”. Extant animals and plants had been classified starting with Linnaeus, and evolution was inferred by comparing traits of organisms. Fossils of ancient animals added to the idea of evolution in deep time. In 1924, Oparin, and later in 1929, Haldane, suggested that a primordial soup would accumulate in a sterile ocean, due to the formation of organic molecules from reduced gasses and energy. This would be the milieu for life to emerge.
With the Miller-Urey experiment (1952) that demonstrated that amino acids, the “basic building blocks of life” could be created quickly in the lab with a primordial atmosphere gas mixture and electricity, it was assumed that proteins that form the basis of most of life’s structure and function would follow. The time needed for the evolution of life was increased from less than 10,000 years in the Biblical Old Testament, to 100 million years (my) in the late 19th century, to about 4.5 billion years (Ga) once radioisotopic dating was established by 1953. Fossil evidence relied on the mineralization of hard structures which started to appear in the Cambrian period around 550 million years ago (mya).
The Apollo lunar samples indicated that the Moon had been subjected to a late heavy impactor bombardment (LHB) after its formation 4.5 Ga from around 4.1 – 3.8 Ga. With the Earth assumed to be sterilized by the LHB, there seemed to be plenty of time for life to appear. Then the dating of stromatolites pushed the earliest known life to nearly 3.5 Ga and reduced the time for abiogenesis to just a few 100 million years after the LHB. This seemed to leave too little time for abiogenesis. There was a reprieve when it was argued that the LHB was an artifact of lunar sample collection, with the later Imbrium impact adding its younger age to the older samples. If the LHB was not a sterilizing event, then another 500 million years to a billion years could be allowed for life to appear.
Even though the structure of DNA was determined by Watson and Crick in 1953, and with it the site of genes, sequencing even short lengths of DNA was a slow process. This changed with gene sequencing machines and algorithms during the 1990s with the sequencing of the human genome. Sequencing costs have fallen sharply, and gene databases are being filled. We now have vast numbers of sequenced genes from a range of organisms, and full genomes from selected species.
The resulting inexpensive gene sequencing kickstarted the genomics revolution. With gene sequences from a large number of extant species, Richard Dawkins suggested that even if there were no fossils, evolution could be inferred by the changes in the nucleotide base sequences in modern organisms, and evolution was represented by the incremental changes in species’ genomes. His opus magnum The Ancestors’ Tale was an exploration of the tree of life moving backwards in time. [6].
The slow changes over time in the sequences of key functional genes that appear in all organisms is called the “molecular clock”. The greater the difference in sequences between the genes in 2 species, the greater their evolutionary separation. However, unlike atomic clocks, the molecular clock does not tick at the same rate for each organism, or gene. If they did, all the divergences would sum to the same length of time. As Figure 1 demonstrates, they do not. Nevertheless, evolutionary trees for all organisms with sequenced conservative functional genes were built to show how species evolved from each other and could be compared with phylogeny trees created using the fossil record.
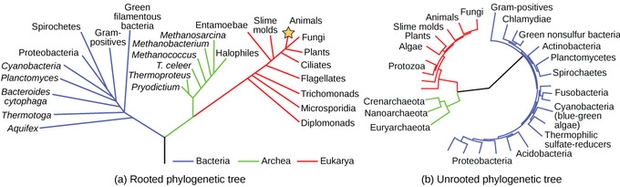
Figure 1. Rooted and unrooted phylogenetic trees. (Source: Creative Commons Chiswick Chap).
While this phylogenetic tree shows evolutionary separation, it has no timeline. These trees converge back in time to a Last Universal Common Ancestor (LUCA) at the point where the 2 most distantly related domains of life, the Bacteria, and Archaea are joined. However, fossils can provide a means to calibrate the timeline for the tree branches and when LUCA can be placed in time. For example, if we can find and date human fossils and chimpanzee fossils, we can be confident that their common ancestor lived at an earlier age. The common ancestor would be younger than the time that both humans and chimps diverged from our ape ancestor, and in turn that ancestor would be younger than the ancestor of all primates. The phylogenetic trees based on gene sequences can be compared to trees based on morphology. Generally, they match. With fossil evidence, these new phylogenetic trees can be calibrated to date the branches.
Without good fossil evidence to calibrate the phylogenetic tree, it is harder to date the tree of life as we approach its root where we believe LUCA must be present. Several attempts have been made to determine this timeline. In 2018, a paper by Betts indicated that LUCA could be dated to about the age of the Earth [2]. Mahendrarajah et al, analyzing the gene for ATP Synthase, estimated a similarly early date for its appearance before the separation of the Archaea and Bacteria placing LUCA at over 4 Ga.[3]
The new paper by Moody et al, extends the work of the aforementioned 2 co-authors, as well as others, to create the best estimate of the timeline of life, the dating of LUCA, a description of LUCA, and its environment. The approach used a cross-bracing method using gene duplications of ancient functional genes to firm up the phylogenetic tree and the fossil calibrations. Cross-bracing is the use of duplicated genes (paralogs) to anchor different trees with dates to provide mutual support for the dating [12].
The 2 different trees are based on gene duplication before LUCA appeared to create the separate trees, which are shown in Figure 2. The analysis dates LUCA at least 4 Ga to the age of the Earth, 4.5 Ga. As most theories of abiogenesis require a watery environment, the earliest dating of surface water on Earth and the appearance of oceans is fairly fast, within 100 million years (my) after Earth’s formation, about 4.4 Ga, [11]. The relaxed Bayesian distributions used hard (no 2.5% tail distribution) and soft (include 2.5% tail distribution) dates for the boundary dating calibrations The maximum likelihood for the age of LUCA was set at 4.2 Ga, 200 my after the oceans were formed and about 300 my after the Earth formed and the impact that formed the moon and sterilized the Earth.
Figure 2 shows the new timeline. The dendrogram indicates the degree of gene sequence divergence as a horizontal line from each node. The greater the length of the line, the more ticks of the molecular clock as the sequence changes compared to nearby species’ lines, and the greater the time the species have been separated by evolution. LUCA is dated within the Hadean eon, a time once thought to be devoid of life due to its hellish surface conditions from impactor bombardment as well as the heat from its formation and radioactivity. The 4.5 Ga calibration date is a hard constraint as terrestrial abiogenesis is impossible before then.
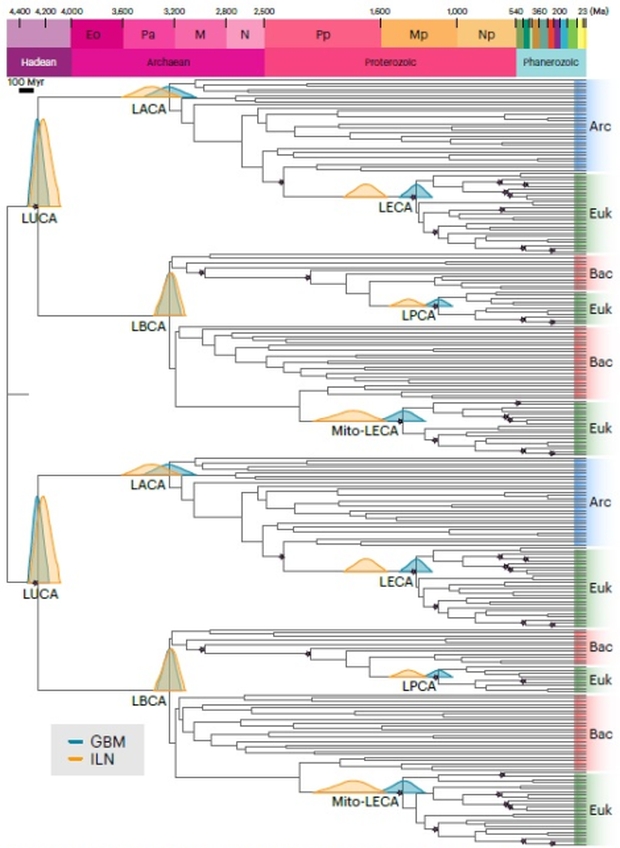
Figure 2. The calibrated phylogenetic tree shows the 2 lineages for the gene duplications, with each of the 2 trees acting as cross braces. The 2 algorithm variants with distributions in gold and teal converge to close overlaps with the dating of LUCA. Note the small purple stars that are the fossil calibrations. The calibrations for LUCA use the age of the Earth and prior fossil evidence as there is no fossil evidence for LUCA unless the controversial carbon isotope evidence demonstrates life and not an abiotic process. Credit: Moody et al.
The paper also uses the gene sequence evidence to paint a picture of LUCA as very similar to a prokaryote bacterium. It has all the important cellular machinery of a contemporary bacterium but with several cellular pathways absent or of low probability. It was probably a chemoautotroph, meaning that it could use free hydrogen and carbon dioxide to reduce and fix carbon as well as extract energy, from either geochemical processes or other contemporary organisms.
Because LUCA is not a protocell, but a likely procaryote, this implies that the sequence of abiogenesis from inanimate chemistry to a functioning prokaryote cell must have taken no more than 300 my, and more likely 200 my.
As the authors state:
How evolution proceeded from the origin of life to early communities at the time of LUCA remains an open question, but the inferred age of LUCA (~4.2 Ga) compared with the origin of the Earth and Moon suggests that the process required a surprisingly short interval of geologic time. (emphasis mine).
The issue of the rapid appearance of life was back in play.
Figure 3 shows the hypothetical progression of abiogenesis to the Tree of Life and the steps needed to get from a habitable world to LUCA at the base of the Tree of Life.
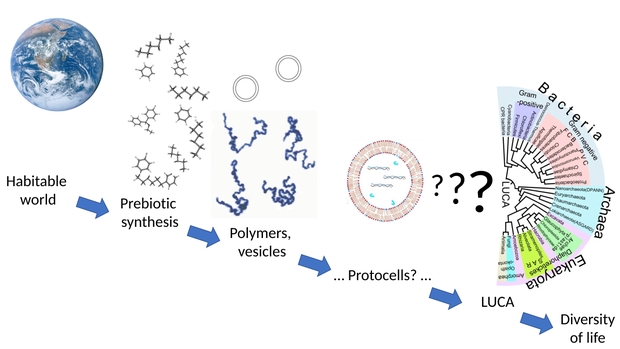
Figure 3. The hypothetical development of life from the habitable planet through simpler stages and eventually to the radiation of species we see today. (Source: Creative Commons Chiswick Chap).
Given that the complexity of LUCA appears to be great, why is the timeline to evolve it so short when the timeline to the last archaean and last bacterial common ancestors (LACA, LBCA) is so prolonged at a billion years? Are the genomic divergences between bacteria and archaea so great not because of a slow ticking of the molecular clock, but rather evidence of rapid evolution that would imply LUCA was younger than it appears as the molecular clock was ticking faster?
It is important to understand that LUCA was not a single organism, but a representative of a population. It probably lived in an ecosystem with other organisms, none of whose lineages survived. This is shown below in Figure 4. The red lines indicating that other extinct lineages may have transferred genes to each of the archaean and bacterial lineages after LUCA evolved could, in principle, have exaggerated the divergence of these 2 lineages, exaggerating the depth of the timeline from LUCA. This is purely speculative to explain the authors’ findings.
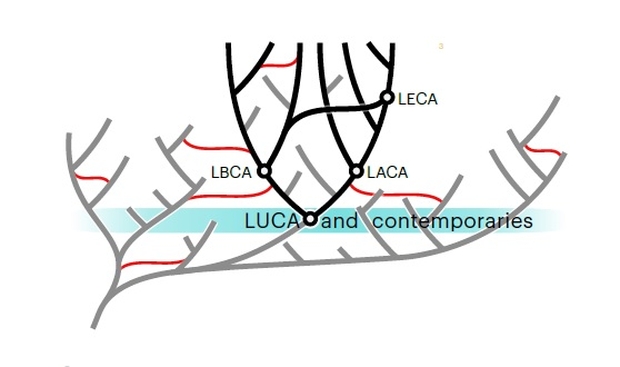
Figure 4. LUCA must have had ancestors and likely contemporary organisms. The gray lineage includes LUCA’s ancestors as well as other lineages that became extinct. The red lines indicate horizontal gene transfer across lineages.
A key question is whether the calibrated timeline is correct. While the authority of the number of authors is impressive, and the many checks on their analysis are substantial, the method may be simply inaccurate. We have a similar methodological issue with the Hubble Tension between 2 methods of determining the Hubble constant for the universe’s rate of expansion. Molecular clock rates are not uniform between species and estimated timelines for the divergence of species can vary when compared to the oldest fossils. DNA sequences can be extracted for relatively recent fossils to more accurately calibrate the phylogenetic tree. However, this is not possible after a few million years due to DNA degradation. Purely mineralized fossils, impressions in rocks, and isotopic biosignature evidence rule this tight calibration out. Fossils are relatively rare and usually prove younger than the node that starts their particular lineage. This is to be expected, although the discovery of older fossils can modify the picture.
Because molecular clock rates are not fixed, various means are used to estimate rates, using Bayesian probability. These rely on different distributions. The authors use 2 methods:
1. Geometric Brownian motion (GBM)
2. Independent log-normal.(ILN)
In Figure 2, the distributions are indicated by color. For the younger nodes, these methods clearly diverge, and in the case of the last eukarya common ancestor, the 2 distributions do not overlap. The distributions converge deeper in time, with the GBM maximum probability now a little older than the ILN one. The authors selected the GBM peak as the best dating for LUCA, although using the ILN method makes almost no difference.
While the Bayesian method has become the standard method for calibrated phylogenetic tree dating, the question remains whether it is accurate. All the genes and cross-bracing used would be false support if there is a flaw in the methodology. A 2023 paper by Budd et al highlights the problem. In particular, based on fossils, the divergence of mammals occurs after the K-T event that is associated with the extinction of the non-avian dinosaurs, whereas the genomic data supports a much older divergence without any fossil evidence. The paper argues that the same applies to the emergence of animals. Fossils in the Cambrian era are much younger than the calibrated phylogenetic data suggests.
Budd states that:
Overall, the clear implication is that the molecular part of the analysis does not allow us to distinguish between different times of origin of the clade, and thus does not contradict the general picture provided by the fossil record….
…we believe that our results must cast severe doubt on all relaxed clock outcomes that substantially predate well-established fossil records, including those affected by mass extinctions.
This becomes extremely problematic when there are no fossils to compare with. In the Moody paper the LACA and LBCA nodes have no calibrations at all, and LUCA has somewhat ad hoc calibration points. If Budd is correct, and he makes a good case, then all the careful analyses of the Moody paper are ineffective, due to fundamental flaws in the tools.
Given the paucity of hard fossil evidence, the known issues of calibrated Bayesian priors for molecular clock dating of phylogenetic trees, compared to the careful testing by the authors of the LUCA paper, the best we can do is look at the consequences of the paper being an over/underestimate of the age of LUCA.
The easy consequence is that the age of LUCA has been overestimated. That LUCA was represented by a population between 3.4 and 4 Gya, with a peak probability somewhere in between. This would allow up to a billion years for abiogenesis to reach this point before the various taxons of archaea and bacteria separated 100s of millions of years later, and subsequently, the eukarya separated from the archaea even later.
This would grant a comfortable period to postulate that at least one abiogenesis happened on Earth and that all life on Earth is local. Conventional ideas on the likely sequence of events remain reasonably intact. Other planets may have their abiogenesis events, with any possibility of panspermia increasingly unlikely with distance. For example, any life discovered in the Enceladan ocean would be a local event with a biology different from Earth’s.
The harder consequences are assuming the short timeline for abiogenesis is correct. What are the implications?
First, it strengthens the argument that under the right conditions, life emerges very quickly. While we do not know what those conditions are exactly, it does suggest that our neighbor, Mars, which has evidence of surface water as lakes and a boreal sea, could have also spawned life. As Mars was not formed after an early collision, its water bodies may date another 100 my before the oceans on Earth. As Mars’ gravity is lower than on Earth, the transfer of material containing any life might have seeded Earth with life.
If we find life in the subsurface of Mars’s crust, it would be important to determine if its biology was the same or different from Earth’s life. If different, that would be the most exciting result as it would argue for the ease of abiogenesis. If the same, then a possible common origin. The same applies to any life that might be found in the subsurface oceans of the icy moons of the outer planets. Different origins imply abiogenesis is common. Astrobiologist Nathalie Cabrol seems quite optimistic about possible life on Mars, and any [dwarf] planet with a subsurface ocean [8]. Radiogenic heating can also ensure liquid water on planets that are well outside the traditional habitable zone (HZ) [10].
If abiogenesis is common, then we should detect biosignatures in many exoplanets in the HZ with the conditions we expect for life to start and thrive. Carr has suggested, rather controversially, that Mars was the better environment for abiogenesis, and therefore terrestrial life was due to panspermia from Mars [5].
What if the rest of the solar system is sterile, with no sign of either extant or extinct life? This would imply the conditions on Earth suitable for abiogenesis are narrower than we thought, which would suggest exoplanet biosignatures would be rarer than we might expect from the detected conditions on those worlds.
The last option is one we would prefer not to be the case if the aim is to work on how abiogenesis occurred on Earth. This option is to accept that LUCA appeared after just a few hundred million years, but that this time was too short. It would imply that the location of abiogenesis, however it occurred, was not on Earth. It would imply that the same probably applies to other bodies in the solar system and therefore life originated in another star system.
Leslie Orgel and Francis Crick’s early suggestion was that terrestrial life was spawned by panspermia [4]. Would that derail studies on the origin of life, or assume only plausible terrestrial conditions? How would we determine the truth of panspermia? I think it could only be demonstrated by sampling life on exoplanets and determining they all shared the same biology fairly exactly. The consequences of that might be profound.
A last thought, that surprised me in my thinking about abiogenesis being seemingly impossibly short: Cabrol, states, with no supporting evidence that [9]:
…how much time it takes for the building blocks of life to transition to biology.….estimates range between 10 million years and as little as a few thousand years.
If true, then life could appear anywhere with suitable conditions, however transient those conditions are. What state that life would be in, for example, protocells, or some state prior to LUCA is not explained [but see Figure 3], but if correct, appears to offer more time for LUCA to evolve. That is indeed food for thought.
References
Moody, E. R. R., Álvarez-Carretero, S., Mahendrarajah, T. A., Clark, J. W., Betts, H. C. Dombrowski, N., Szánthó, L. L., Boyle, R. A., Daines, S., Chen, X., Lane, N., Yang, Z., Shields, G. A., Szöllősi, G. J., Spang, A., Pisani, D., Williams, T. A., Lenton, T. M., & Donoghue, P. C. J. (2024). The nature of the last universal common ancestor and its impact on the early Earth system. Nature Ecology & Evolution. https://doi.org/10.1038/s41559-024-02461-1 https://www.nature.com/articles/s41559-024-02461-1
Betts, H. C., Puttick, M. N., Clark, J. W., Williams, T. A., Donoghue, P. C. J., & Pisani, D. (2018). Integrated genomic and fossil evidence illuminates life’s early evolution and eukaryote origin. Nature Ecology & Evolution, 2(10), 1556–1562. https://doi.org/10.1038/s41559-018-0644-x https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6152910/
Mahendrarajah, T. A., Moody, E. R. R., Schrempf, D., Szánthó, L. L., Dombrowski, N., Davín, A. A., Pisani, D., Donoghue, P. C. J., Szöllősi, G. J., Williams, T. A., & Spang, A. (2023). ATP synthase evolution on a cross-braced dated tree of life. Nature Communications, 14(1). https://doi.org/10.1038/s41467-023-42924-w
F.H.C. Crick, L.E. Orgel, (1973) “Directed panspermia”, Icarus, Volume 19, Issue 3, Pages 341-346, ISSN 0019-1035, https://doi.org/10.1016/0019-1035(73)90110-3.
Carr, C. E. (2022). Resolving the history of life on Earth by seeking life as we know it on Mars. Astrobiology, 22(7), 880–888. https://doi.org/10.1089/ast.2021.0043 https://arxiv.org/pdf/2102.02362
Dawkins, R. (2004). The Ancestor’s Tale: A Pilgrimage to the Dawn of Evolution. Houghton Mifflin Harcourt.
Budd, G. E., & Mann, R. P. (2023). Two notorious nodes: a critical examination of relaxed molecular clock age estimates of the bilaterian animals and placental mammals. Systematic Biology. https://doi.org/10.1093/sysbio/syad057
Cabrol, N. A. (2024). The secret life of the universe: An Astrobiologist’s Search for the Origins and Frontiers of Life. Simon and Schuster.
Ibid. 148.
Tolley, A (2021) “Radiolytic H2: Powering Subsurface Biospheres” https://www.centauri-dreams.org/2021/07/02/radiolytic-h2-powering-subsurface-biospheres/
Elkins-Tanton, L. T. (2010). Formation of early water oceans on rocky planets. Astrophysics and Space Science, 332(2), 359–364. https://doi.org/10.1007/s10509-010-0535-3
Sharma, P. P., & Wheeler, W. C. (2014). Cross-bracing uncalibrated nodes in molecular dating improves congruence of fossil and molecular age estimates. Frontiers in Zoology, 11(1). https://doi.org/10.1186/s12983-014-0057-x
Background Reading
The Hadean-Archaean Environment
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2869525/
History of Earth
https://en.m.wikipedia.org/wiki/History_of_Earth
Hadean
https://en.m.wikipedia.org/wiki/Hadean
Late Heavy Bombardment
https://en.m.wikipedia.org/wiki/Late_Heavy_Bombardment
Wikipedia: Portal: Evolutionary Biology
https://en.wikipedia.org/wiki/Portal:Evolutionary_biology
Origin of life: Drawing the big picture
https://www.sciencedirect.com/science/article/abs/pii/S0079610723000391
The Origin of Life: What We Do and Don’t Know
https://hea-www.harvard.edu/lifeandthecosmos/wkshop/sep2012/present/CleavesSILifeInTheCosmosTalk2012b.pdf
Introduction to Origins of Life of Earth
https://pressbooks.umn.edu/introbio/chapter/originsintro/
Abiogenesis
https://en.wikipedia.org/wiki/Abiogenesis
Last universal common ancestor
https://en.wikipedia.org/wiki/Last_universal_common_ancestor
Earth’s timeline
https://dynamicEarth.org.uk/geological-timeline-pack-2.pdf
Formation of early water oceans on rocky planets
https://link.springer.com/article/10.1007/s10509-010-0535-3
Earliest known life forms
https://en.wikipedia.org/wiki/Earliest_known_life_forms
Molecular clock
https://en.wikipedia.org/wiki/Molecular_clock
Phylogenetic Tree
https://en.wikipedia.org/wiki/Phylogenetic_tree
Primordial Soup
https://en.wikipedia.org/wiki/Primordial_soup



For those interested in seeing living examples of stromatolites, the oldest dated fossil signs of life (3.5-3.8Gya) and mentioned above, there are several locations that can be visited: Shark Bay, a national park, in western Australia; just off shore in some islands in the Bahamas; Lagoa Salada in Brazil; Pampa de Tamarugal National Reserve in Chile; Coahuila and Bacalar in Mexico and somewhat surprisingly in the Great Salt Lake in Utah. These are colonies of single-celled cyano-bacteria and perhaps other species, mixed with sand and precipitating calcium carbonate, that grow slowly over time. The living versions are not very colorful. Some fossils of them, on the other hand, have mineralized very colorfully.
Excellent Analysis, Alex!
My own two cents would be
1) You seem to be citing Budd who explains how the short timeline may be an illusion, but then go on assuming it to be true anyway. I would be far more skeptical of the early molecular clock calibrations, in agreement with Budd.
2) Even the “short” timeline of hundreds of Ma is not really short at all.
@Eniac
Conventional “wisdom” was that getting from chemistry to, e.g. bacteria, needed a long time. This may not be true. What is true is that making even more than basic steps to the simplest life has eluded us so far. Progress has been made, but not as much as was once thought would happen by now. Maybe it is life AGI, some experiments will change that POV.
In the event, I have tried to show agnosticism and offer some “predictions” based on different conditions for abiogenesis.
As a biologist, I would definitely prefer a terrestrial location, and that abiogenesis is “easy”, as that would likely mean more life with differing biologies. That would be a goldmine for research into living processes when all we have is one common type on Earth, as far as we know. I would love to be alive if we discover biosignatures, hopefully not too ambiguous, on exoplanets, and even life elsewhere in our system that we could sample and study in the lab.
However, we are not there yet, and it may turn out life is very rare. I await data to tip my POV either way!
Another comment to be made is that the biogenicity of the oldest stromatolites is speculative and should not be considered settled or used as a secure premise for dating purposes. See, eg, here: https://pubmed.ncbi.nlm.nih.gov/11540142/
That paper is from 1994. I don’t believe that there remains any serious doubt that the Strelley Pool rocks do contain fossilized bacterial mats consistent with stromatolites. The ambiguity has shifted to far older isotopic ratios that could be indicative of life or possibly, abiotic processes.
It is true that the paper is somewhat old, but I don’t think that later work has changed the speculative nature of microfossils much. There are abiological explanations for stromatolites, and I could find no paper that made claims that the matter is settled.
@Eniac,
I concede it is not certain the Strelley Pool “stromalites” are indeed stromatolites.
This Google Scholar search reveals both sides of the biotic/abiotic argument.
If they are abiotic, then their use as calibration points is suspect. What I don’t know is how much this would change the model output. When I was researching this article, none of the authors replied to an email requesting the impact of changing the calibration points.
Hi Paul & Alex
This seems to strongly hint that LUCA came to Earth via Panspermia. Thus the Origin of Life could well be pushed back closer and closer to the Big Bang. Loeb has suggested the very early days of the cosmic expansion provided habitats warmed via the CMB.
@Adam
I just wanted to cover the bases and allow for panspermia as an option. In my mind, panspermia covers transfer by interplanetary impactors, such as from Mars to Earth, stellar winds blowing bacterial spores out from a living world to the outer planets and beyond, interstellar visitors like Borisov that are refuges for bacteria [spores], and accidental transfer by visiting spacecraft, and lastly by directed panspermia to seed exoplanets deliberately.
To use the “fine-tuning” argument of Kipping and Lewis, the more difficult abiogenesis is, the more likely that panspermia from existing life is the mechanism. In our living world, life no longer spontaneously appears, it is entirely due to the spread of existing life through various means, and why sterilization is so important for some settings.
If we are wrong about the time needed to evolve a “minimal prokaryote” and it is very short rather than long, the abiogenesis is favored, assuming the conditions are favorable. Recall my post concerning the paper about which amino acids seem to be energetically favored in mafic basalts. (I think that discovery could be important as it appears associated with particular protein structures – alpha helices – that seem more prevalent in the organisms using H2 to reduce carbon for growth. – e.g. in methanogen archaea. Energetics of Archaean Life in the Ocean Vents (2022).)
Wow Alex
That was a very interesting and informative read, great graphics there too.
Plenty to ponder
Thanks Edwin
This is a very helpful overview that helps us understand the ongoing controversies. What we should make of it seems to hinge on statistical thinking that is not easy to do. I was very excited to think that Bacteria and Archaea could have been separate colonizations from an external source. But it’s also possible that the unique conditions of the Hadean, rather than being harmful to life, were vitally important for abiogenesis. For example, https://link.springer.com/article/10.1186/1467-4866-15-8 describes some ways in which pyrophosphate and polyphosphates might be created geologically, and have been observed in fumaroles in Japan – while observing that these chemicals are very rare on modern Earth. Polyphosphates are like “free ATP” for living organisms, a readily tapped energy source for RNA-based life. Perhaps a planet gets one good shot at converting an era of widespread volcanic activity into metabolic energy.
@ MS
To strain a metaphor, abiogenesis may be like baking. It may be relatively easy like Banana Bread which tolerates very wide ranges of ingredients, yet still seems to work. OTOH, if you have ever watched the British “Bake Off” TV series, each show has a technical challenge that requires quite precise use of ingredients and baking conditions. We just don’t know about abiogenesis. What ingredients do we need? Is there something missing (like baking powder or soda with cake and bread) that prevents abiogenesis from working? Or do the conditions need to be exactly right? Does the incorporation of bacteria into an Archaean need a 1:1bn exact condition to start the ball rolling for Eukarya, or is this a nearly inevitable outcome? It may be that all our reductionist experiments are failing to complete the needed recipe.
I certainly agree… the trick is trying to guess the missing ingredient. There are so many possibilities! One I was wondering about in this context is Theia. We normally think of the Theian collision as favored by the anthropic principle for giving us a Moon that keeps our axial tilt and climate constant, but could it have had an effect on abiogenesis? Is it possible the combined planet could have been left with one side Hadean, one side ‘terraformed’, for an extended period of time? It takes volcanic circumstances and acidic conditions to convert phosphate rock into polyphosphates, but the polyphosphates can only be used for energy in a nice aqueous primordial soup, in which they’ll break down spontaneously in no time.
Do the 2 conditions have to be simultaneous? If the Theia collision created conditions to create polyphosphate, could those compounds remain available for a later period when the oceans formed? Alternatively, could not both conditions have existed together, with polyphosphate created after the impact yet quickly dissolved in transient, condensed hot water pools before conditions settled down to allow oceans to form?
As we don’t know how life started, we may be focused too much on terrestrial life. It is the same problem as assuming ETI must be embodied in humanoid forms and all the chance events that led to us. Maybe life has a variety of possible biologies that appear contingent on local conditions. To me, that is what makes exploration so exciting, as any discovery of extraterrestrial life would give us information on how convergent or divergent biology is.
A hodgepodge of thoughts:
Alex, are you a fan of punctuated equilibrium…
https://en.wikipedia.org/wiki/Punctuated_equilibrium
Making organic soup on Cosmos in 1980…
https://www.youtube.com/watch?v=_2xly_5Ei3U
Surviving for over 100 million years…
https://www.science.org/content/article/scientists-pull-living-microbes-100-million-years-beneath-sea
Or how about two BILLION years old and possibly still alive…
https://www3.nhk.or.jp/nhkworld/en/news/backstories/3472/#:~:text=A%20University%20of%20Tokyo%20research%20team%20says%20it,the%20origin%20and%20evolution%20of%20life%20on%20Earth.
Excellent article. It would seem that if there is living life on Mars, it may be way below the surface of the planet. That doesn’t mean some of it couldn’t find its way upwards.
The Moon may still hold fossil remains from Earth brought there via impacts…
https://www.science.org/content/article/ancient-earth-rock-found-moon
Venus is looking like a better prospect than ever for present life. I wonder if Venus once had oceans and therefore life if any fossils remain? So much of that amazing world we have barely scratched.
The outer Sol system worlds still hold much promise. Frustrating that it is taking so long to start really exploring them. And we need to expand our search to certain planetoids and worlds way beyond the Saturn system.
https://www.planetary.org/articles/meet-orbilander-enceladus-mission
https://ntrs.nasa.gov/api/citations/20210014731/downloads/2021.MacKenzie.EnceladusOrbliander.pdf
@LJK
You have posted a lot of interesting articles I need to read first. However, to respond to “preferring evolution by creeps vs jerks”. This is an old dispute and has surely been relegated to history by abundant work using genetic algorithms on computers. They demonstrate that both rapid changes followed by a long period of stasis appear as well as long periods of slowly increasing changes. It so depends on the fitness landscape and local conditions.
I would hope that Niles Eldredge could admit that both his preferred phyletic gradualism AND Gould’s preferred punctuated equilibrium are valid models, dependent on conditions. While Gould is no longer with us, both lived to see computer modeling with GAs demonstrate both forms, at least genomically and in fitness changes.
I would say that phyletic gradualism if reflecting the molecular clock is the preferred model for phylogenetic trees as used in this paper, but that doesn’t mean that the clock must tick at an even rate, just appears to do so when averaged over a long time.
BTW, geneticists long knew of the different evolutionary rates of large versus small populations, founder effects, etc., long before the molecules carrying those genes were known.
1. 2 bn year old life.
I would be very interested in any analysis which is published. Please keep an eye out for any. This is hard data compared to molecular clock extrapolations. I don’t expect it, but imagine if they find sequences that cannot be linked to the existing phylogenetic tree.
2. Ancient Earth rock found on the moon
The article is 5 years old. I haven’t read anything about further discoveries. It could well be a lucky find as we only have the relative paucity of Apollo rocks to work with.
3. Orbilander
I am pleased that the concept has included a microscope. The nanopore DNA sequencer is small, so no great loss if any life doesn’t have DNA as its information storage. But if we discovered “wiggling bugs” but they didn’t have DNA, that would imply a different biology and probable different abiogenesis.
4. Oxygen Produced In The Deep Sea Raises Questions About Extraterrestrial Life
I am still looking for information on how much difference this makes to the O2 concentration in the deep ocean. As it is towards the end of the path of global ocean circulation, I would expect that it would reverse the slow decline of O2, But does it?
5. Origins of Life: The Protein Folding Problem All Over Again?
A very interesting paper that I want to study more carefully. I am not sure I agree with the premise based on a first quick read, but it certainly bears study.
It is a pity I don’t have access to any reviewers to gain insights from any of their observations.
Punctuated equilibrium sounds very mysterious on a nature documentary, but it is part of your mundane life. One year you lived in Illinois and you drove from an apartment to a job site and various nearby eateries and the occasional tourist attraction. Then all of a sudden you were driving around Vermont all the time. What happened? Well, the “car” (the rate evolution can change the physical form of an organism) can move very fast, but the place it has to go to survive (“work”, its ecological niche) may stay the same for a long time. But then it finds another niche, and it’s gone across the country in a flash.
https://astrobiology.com/2024/08/uncovering-the-role-of-oxygen-concentration-in-the-formation-of-early-earth-magma-ocean.html
Uncovering The Role Of Oxygen Concentration In The Formation Of Early Earth Magma Ocean
By Keith Cowing
Press Release
Okayama University
August 28, 2024
It is widely accepted that the early Earth largely consisted of molten magma, forming a global ocean of magma. This extreme state of Earth was likely caused by the intense heat generated from accretionary impacts, meaning the collision of smaller celestial bodies with Earth.
Understanding the formation of this magma ocean is crucial for comprehending Earth’s formation. A major problem with current magma ocean formation models is the lack of consensus on the melting temperatures of deep mantle rocks. Models explaining Earth’s core formation use a specific set of experimental data to estimate mantle melting temperatures, but recent experiments have shown that these temperatures may differ by 200–250 °C from the previously accepted data.
Some studies indicate that oxygen fugacity, or the amount of oxygen available in the mantle, may strongly affect the melting temperatures of deep mantle rocks, which in turn may have influenced the formation of the magma ocean. The mantle’s oxygen fugacity is thought to have increased during accretion, core formation and subsequent mantle evolution; however, the effect of this increase on the melting temperatures of deep mantle materials remains unclear.
Addressing this gap, a team of researchers led by Associate Professor Takayuki Ishii from the Institute for Planetary Materials at Okayama University, Japan and Dr. Yanhao Lin from the Center for High Pressure Science and Technology Advanced Research, China, investigated the effects of oxygen fugacity on magma ocean formation during early Earth evolution.
“The evolution of early Earth has been greatly influenced by oxygen fugacity, which may necessitate the reconsideration of current models. To this end, we assessed the effect of oxygen fugacity on the melting temperatures of deep mantle materials to constrain the conditions at the floor of a deep terrestrial magma ocean,” explains Prof. Ishii.
The study also involved Professor Wim van Westrenen from the Department of Earth Sciences, Faculty of Science at Vrije Universiteit Amsterdam, the Netherlands, Professor Tomoo Katsura from Bayerisches Geoinstitut, University of Bayreuth, Germany, and Dr. Ho-Kwang Mao from the Center for High Pressure Science and Technology Advanced Research, China. It was published online in the journal Nature Geoscience on July 16, 2024.
The researchers conducted melting experiments at pressures of 16–26 Gigapascals, similar to mantle depths between 470 km and 720 km, at high oxygen fugacities, on mantle pyrolite, a material composition representing Earth’s mantle. Results revealed that over this pressure range, the melting temperatures decreased with increasing oxygen fugacity and were at least 230–450 °C lower than those from experiments conducted at low oxygen fugacities.
Assuming a constant temperature for the magma ocean, this implies that the magma ocean floor deepens by about 60 km for each logarithmic unit increase in mantle oxygen fugacity. This strong influence of oxygen fugacity on mantle melting suggests that current models for early Earth thermal evolution and core formation need re-evaluation.
Furthermore, these results can also explain the apparent discrepancy between the low oxygen fugacities predicted for the Earth’s deep mantle post-core formation and the high oxygen fugacities observed in magmatic rocks over 3 billion years old, formed by melting of the deep mantle.
“Beyond Earth’s formation, our findings on the dependence of melting temperatures on oxygen fugacity can also be applied to understand the formation of other rocky planets that can support human life,” remarks Dr. Lin, highlighting the potential impact of the study. He adds, “For example, these results can improve our understanding of Mars, which is a recent hot topic regarding human habitability.”
This groundbreaking study promises to improve Earth formation models, deepening our understanding of the formation of Earth and other such planets.
About Okayama University, Japan
As one of the leading universities in Japan, Okayama University aims to create and establish a new paradigm for the sustainable development of the world. Okayama University offers a wide range of academic fields, which become the basis of the integrated graduate schools. This not only allows us to conduct the most advanced and up-to-date research, but also provides an enriching educational experience.
Website: https://www.okayama-u.ac.jp/index_e.html
Melting at the base of a terrestrial magma ocean controlled by oxygen fugacity, Nature Geoscience (open access).
https://astrobiology.com/2024/08/a-reassessment-of-the-hard-steps-model-for-the-evolution-of-intelligent-life.html
A Reassessment of the “Hard-steps” Model For The Evolution Of Intelligent Life
By Keith Cowing
Status Report
astro-ph.EP
August 27, 2024
While there exist more contentious geochemical and molecular clock estimates for these steps that would place them each farther back in time, we have chosen the least controversial evidence to produce the most conservative timeline possible. Therefore, each candidate hard step necessarily preceded, but occurred no later than, their displayed dates, and the incorporation of other lines of evidence would necessarily shift the origin of each step back in time to varying degrees.
The time intervals separating adjacent steps were calculated using the minimum age constraints only and are displayed in bold and expressed in billions of years (Gyr). With respect to the eukaryotic fossil record, there is ongoing uncertainty concerning when the last eukaryote common ancestor (LECA) evolved (55), which marks the completion of the ‘eukaryogenesis’ process (51). Specifically, it remains unclear whether the LECA emerged hundreds of millions of years before the oldest eukaryotic-grade fossils (1.63-1.67 billion years ago, or Ga), or hundreds of millions of years after (to use the two end-member scenarios) (55).
In order to explore the implications of both scenarios, we display both: 1) the oldest fossil evidence for recognizable crown-group eukaryotes (1.06-1.03 Ga), which designates all eukaryotes, extant and extinct, descended from the LECA, and 2) the oldest fossil evidence for total-group eukaryotes (1.67-1.63 Ga), which comprises both crown-group eukaryotes and the now-extinct eukaryote lineages that diverged prior to the LECA (stem-group eukaryotes). A cladogram depicting the concepts of total, stem, and crown groups is displayed on the left, with † designating extinct stem-group lineages. Data sources: oldest evidence for life (56–59); oxygenic photosynthesis (60, 61); total-group eukaryotes (62); crown-group eukaryotes (63); crown-group animals (64); and Homo sapiens (65). — q-bio.PE
According to the “hard-steps” model, the origin of humanity required “successful passage through a number of intermediate steps” (so-called “hard” or “critical” steps) that were intrinsically improbable with respect to the total time available for biological evolution on Earth.
This model similarly predicts that technological life analogous to human life on Earth is “exceedingly rare” in the universe.
Here, we critically reevaluate the core assumptions of the hard-steps model in light of recent advances in the Earth and life sciences. Specifically, we advance a potential alternative model where there are no hard steps, and evolutionary novelties (or singularities) required for human origins can be explained via mechanisms outside of intrinsic improbability.
Furthermore, if Earth’s surface environment was initially inhospitable not only to human life, but also to certain key intermediate steps in human evolution (e.g., the origin of eukaryotic cells, multicellular animals), then the “delay” in the appearance of humans can be best explained through the sequential opening of new global environmental windows of habitability over Earth history, with humanity arising relatively quickly once the right conditions were established.
In this co-evolutionary (or geobiological) scenario, humans did not evolve “early” or “late” with respect to the total lifespan of the biosphere, but “on time.”
Daniel B. Mills, Jennifer L. Macalady, Adam Frank, Jason T. Wright
Subjects: Populations and Evolution (q-bio.PE); Earth and Planetary Astrophysics (astro-ph.EP)
Cite as: arXiv:2408.10293 [q-bio.PE] (or arXiv:2408.10293v1 [q-bio.PE] for this version)
https://doi.org/10.48550/arXiv.2408.10293
Submission history
From: Daniel Mills
[v1] Mon, 19 Aug 2024 16:30:06 UTC (1,829 KB)
https://arxiv.org/abs/2408.102933
https://astrobiology.com/2024/08/oxygen-produced-in-the-deep-sea-raises-questions-about-extraterrestrial-life.html
Oxygen Produced In The Deep Sea Raises Questions About Extraterrestrial Life
By Keith Cowing
Press Release
Boston University
August 27, 2024
Over 12,000 feet below the surface of the sea, in a region of the Pacific Ocean known as the Clarion-Clipperton Zone (CCZ), million-year-old rocks cover the seafloor. These rocks may seem lifeless, but nestled between the nooks and crannies on their surfaces, tiny sea creatures and microbes make their home, many uniquely adapted to life in the dark.
These deep-sea rocks, called polymetallic nodules, don’t only host a surprising number of sea critters. A team of scientists that includes Boston University experts has discovered they also produce oxygen on the seafloor.
The discovery is a surprise considering oxygen is typically created by plants and organisms with help from the sun—not by rocks on the ocean floor. About half of all the oxygen we breathe is made near the surface of the ocean by phytoplankton that photosynthesize just like land-dwelling plants.
Since the sun is needed to carry out photosynthesis, finding oxygen production at the bottom of the sea, where there is no light, flips conventional wisdom on its head. It was so unexpected that scientists involved in the study first thought it was a mistake.
“This was really weird, because no one had ever seen it before,” says Jeffrey Marlow, a BU College of Arts & Sciences assistant professor of biology and coauthor on the study, which was published in Nature Geoscience.
As an expert in microbes that live in the most extreme habitats on Earth—like hardened lava and deep-sea hydrothermal vents—Marlow initially suspected that microbial activity could be responsible for making oxygen. The research team used deep-sea chambers that land on the seafloor and enclose the seawater, sediment, polymetallic nodules, and living organisms. They then measured how oxygen levels changed in the chambers over 48 hours.
If there are plentiful organisms breathing oxygen, then the levels would normally decline, depending on how much animal activity is present in the chamber. But in this case, oxygen was increasing.
“We did a lot of troubleshooting and found that the oxygen levels increased many more times following that initial measurement,” Marlow says. “So we’re now convinced it’s a real signal.”
He and his colleagues were aboard a research vessel tasked with learning more about the ecology of the CCZ, which spans 1.7 million square miles between Hawaii and Mexico, for an environmental survey sponsored by The Metals Company, a deep-sea mining firm interested in extracting the rocks en masse for metals.
After running experiments on board the vessel, Marlow and the team, led by Andrew Sweetman at the Scottish Association for Marine Science, concluded the phenomenon isn’t primarily caused by microbial activity, despite the abundance of many different types of microbes both on and inside the rocks.
Polymetallic nodules are made of rare metals, including copper, nickel, cobalt, iron, and manganese, which is why companies are interested in mining them. It turns out, according to the study, that those densely packed metals are likely triggering “seawater electrolysis.” This means that metal ions in the rock layers are distributed unevenly, creating a separation of electrical charges—just like what happens inside of a battery.
This phenomenon creates enough energy to split water molecules into oxygen and hydrogen. They named this “dark oxygen,” since it’s oxygen made with no sunlight. What remains unclear is the exact mechanism of how this happens, if oxygen levels vary across the CCZ, and if the oxygen plays a significant role in sustaining the local ecosystem.
The Metals Company calls polymetallic nodules a “battery in a rock,” and on its website states that mining them could accelerate the transition to battery-powered electric vehicles and claims mining on land would eventually no longer be necessary. So far, mining in the CCZ is exploratory, but the United Nations International Seabed Authority, which manages the area, could start making decisions about mining as soon as next year.
The Metals Company is working with the Pacific states of Nauru, Tonga, and Kiribati to gain access to mining licenses, but many other nations in the South Pacific, including Palau, Fiji, and Tuvalu, have vocally supported a moratorium or a pause to mining plans. Environmental activist groups like Greenpeace and Ocean Conservancy are calling for a permanent ban, and opponents of the operation fear it could cause irreversible damage to the seafloor.
In the meantime, scientists have begun studying the potential impacts of disturbing a largely unexplored ecosystem. This Nature Geoscience paper contributes insights about the baseline conditions of the area before any large-scale mining begins.
“We don’t know the full implications, but to me this finding suggests that we should deeply consider what altering these systems would do to the animal community,” Marlow says, since all animals need oxygen to survive.
The CCZ is also the perfect environment to study the planet’s smallest organisms, such as bacteria and archaea (single-celled organisms) found in sediments and on the nodules. Marlow and his coauthor Peter Schroedl (GRS’25), a PhD student in BU’s ecology, behavior, and evolution program, are especially focused on using microbes found in extreme environments like the CCZ as templates for finding single-celled life on other planets and moons—since deserts, volcanoes, and seafloor vents are the most similar places we have to Mars and Saturn’s many moons. This is called astrobiology, a field that seeks to inform the search for extraterrestrial life by studying Earth’s systems.
“Life in environments like the CCZ provides an opportunity to study ecosystems that developed under distinct evolutionary pressures and constraints,” says Schroedl, who works in Marlow’s lab. Those conditions—the depth, pressure, and aquatic environment—are “analogous to conditions we have measured or expect to discover on icy moons,” he says.
For example, Jupiter’s moon, Enceladus, and Saturn’s moon, Europa, are covered with layers of ice with no sunlight reaching that water trapped underneath. “Who knows—if these types of rocks are under the ice making oxygen, that could allow a more productive biosphere to exist,” Marlow says. “If photosynthesis isn’t required to make oxygen, then other planets with oceans and metal-rich rocks like these nodules could sustain a more evolved biosphere than we’ve thought possible in the past.”
There are plenty of questions to continue asking, Marlow says, about what the dark oxygen discovery means for extraterrestrial oceans and our own.
“For the most part, we think of the deep sea as a place where decaying material falls down and animals eat the remnants. But this finding is recalibrating that dynamic,” he says. “It helps us to see the deep ocean as a place of production, similar to what we have found with methane seeps and hydrothermal vents that create oases for marine animals and microbes. I think it’s a fun inversion of how we tend to think about the deep sea.”
Republishers are kindly reminded to uphold journalistic integrity by providing proper crediting, including a direct link back to the original source URL here:
https://www.bu.edu/articles/2024/deep-sea-oxygen-raises-questions-about-extraterrestrial-life/
Evidence of dark oxygen production at the abyssal seafloor, Nature Geoscience (open access):
https://www.nature.com/articles/s41561-024-01480-8
https://astrobiology.com/2024/08/origins-of-life-the-protein-folding-problem-all-over-again.html
Origins Of life: The Protein Folding Problem All Over Again?
By Keith Cowing
Status Report
PNAS
August 27, 2024
How did specific useful protein sequences arise from simpler molecules at the origin of life? This seemingly needle-in-a-haystack problem has remarkably close resemblance to the old Protein Folding Problem, for which the solution is now known from statistical physics.
Based on the logic that Origins must have come only after there was an operative evolution mechanism-which selects on phenotype, not genotype-we give a perspective that proteins and their folding processes are likely to have been the primary driver of the early stages of the origin of life.
Origins of life: The Protein Folding Problem all over again?, PNAS (open access):
https://www.pnas.org/doi/10.1073/pnas.2315000121
A.T.,
Many thanks for collecting all this together in a concise readable survey.
It arrives at a time when I can well appreciate it. As an office and house cleaning measure, I started out sifting through decades of Science weeklies. And then became struck by all the articles about such milestones that your current article touches on. Most notably from the exercise, free oxygen was a development that started 500 millioon years ago and ancestors like the Edicarans appeared subsequent to that. [And please pardon if I go on repeating what you said above below. But it is a great framework.]
As was noted by A. C. above, what with the harshness of the terrestrial environment for eons in our terms, it could not have been much rougher for our ancestors not to be inhabiting the Earth at all. Dropping from a circumsolar disk onto a surface like the Earth would be perhaps the roughest part. Until the lunar incident.
In addition, when we speak of oceans, the picture that would have come to my mind and many others for much of life would be that of when one lives by or drives out to its shore. But for oceans to exist on Earth, we have indirectly come to understand that there must be oceans beneath our feet as well. Internal heating drives terrestrial waters to the surface. Or else cooling allows seepage back down. And several icy or terrestrial objects in the solar system seem to uphold that model: Mars and Galilean moons. As for Venus, maybe someone can provide a current or historical model. But depleted no doubt.
But I could walk away at this point relatively convinced of an ancient life scenario dating back to Earth formation except for the circumstances of the forming of the moon. How could anything but INORGANIC chemistry survive such a collision?
Wouldn’t the Earth have a high effective surface temperature for millennia or mega-years to come? The moon’s origin is better explained than what the Earth was like before and after.
Although we do have environments on Earth where ambient oxygen is not the breath of life, which allows bacteria ( if not archaea) to integrate into the biosphere, it is not until about 500 million years ago that oxygen levels rise in the atmosphere and allow for the development of phyla with which we have become familiar and eventually creatures like ourselves. Much of our hopes about getting in contact with the “Where is everybody” crowd rests on the idea that this is a duplicatable path. Evidence such as you collected suggests that “life basic” has what it takes to survive, but it is unclear why it should unfold much further than it had after an eon. And should it have done so elsewhere, why it should resemble anything we have encountered in the geological record thus far.
Well, for us readers it is now a better defined problem with a very detailed “straw man”.
Thanks again.
Thank you Alex for this article. Do you think that the interstellar environment (solar radiation, gamma etc.) play a role in abiogenesis?
Thank you, Alex, for this fine and thought-provoking discussion.
One could think of biological life as a complex molecular machine consisting of many modules that mutually interact to regulate each other to produce the final result. Unlike a physical machine with physical cogs that do not interact with remote regions of the machine, biological cogs and modules may be diffusely located and have widespread interactions.
Likewise a hypothesized ancient organism may have been an array of various functions spread across a community of related organisms and which survived the weeding out and congregated into one or a few entities, prior to multicellularity.
And even after multicellularity as in insect endosymbionts.
And even in placental mammals, endogenization of retroviruses.
To LUKA – an ode?
Spontaneous generation of life always seems to be lurking in the background in discussions such as this. And one might say that this instance does not really refute the concept, but perhaps pushes it back to before the formation of the Earth; that is, if LUKA really started out in a stellar nursery nebula or elsewhere.
Once endorsed by Aristotle, as a contemporary event, spontaneous generation came under scientific scrutiny in the 17th century. Maggots were widely thought to be “spontaneous” and, akin to lightning bolts, drawn to rotting meat. But Francesco Redi in 1688 demonstrated how flies were their agents of distribution with sealed and open containers of meat manifesting none and some respectively.
Now how did Aristotle weigh in on all of this?
“Animals and plants come into being in earth and in liquid because there is water in earth, and air in water, and in all air is vital heat so that in a sense all things are full of soul. Therefore living things form quickly whenever this air and vital heat are enclosed in anything. When they are so enclosed, the corporeal liquids being heated, there arises as it were a frothy bubble.”
— Aristotle, Generation of Animals, Book III, Part 11 (wikipedia translation)
His arguments are not necessarily entirely invalid, but modified by degree or instance. For example, do we assume LUKA is original or our best traceback – to what we don’t know.
If LUKA dropped in on a very different Earth and prospered, then it is possible that LUKA did the same with other solar system bodies. At least the first part. We don’t really know how well it fared. And when the space age arrived, many in the science community were more excited about sampling comets or other bodies for life’s precursors – or even primitive forms. From that “strictly science” viewpoint the biggest bonanza from the moon was the capability to determine its origin – eventually.
So, at this date we still have not been able to verify whether the terrestrial LUKA experience was duplicated elsewhere in the solar system. Largely because the resources for a thorough search have yet to be devised or completed in their operation. Martian landers have not yet been conclusive. Much the same could be said for Galilean moons or those of Saturn.
It is possible that some environments are now more hostile than the supposed environment that in which an ET LUKA might have existed or formed. Contemporary Venus, for example, would be such a case, perhaps the E-M and radiation surrounding Jupiter. Mars, however, is so near to be resolved.
But given the hostility to our own form of life for most of Earth’s history, why should we assume that an early Outer Planet would be out of the question?
If anything, Jupiter is visibly a vast chemical cauldron. The others are more subtly so.
Then, those parts of the solar system which are in deep freeze might also have some trace or evidence of LUKA. We might be falling back to the reasoning that motivated Fred Hoyle and his colleagues when they wrote of viruses on comets.
Their arguments have been looked at skeptically over the years. But orthodoxy has its conundrums too.
The Curiosity and Perseverance rovers are measuring rock samples on Mars at about the same age as LUCA, and the chemistry of the rocks and water bodies from then was oxidizing. You get Manganese in its 4+ state, which is the same state as aerated water on Earth.
Given that early life developed in anaerobic conditions, this would mean that the surface of Mars was inimical to life by that time.
However, this does not mean that the depths of Mars’s Northern Ocean would not have been anaerobic, much like the Black Sea, and so life could have evolved there.
The only way to find this out would to drill through Mars’s northern ice cap and a kilometer or so of rock after that, quite an undertaking. It would however be fruitful as during periods of liquid water on Mars surface soil would have washed into the ocean been deposited on its floor. Rock cores from the ocean floor could give the entire early history of Mars.
In the absence of a continuous overhead canopy permitting brachiation, LUCA or no LUCA, the shoulder would be limited to a single plane of motion as in ungulates, carnivora, and pretty much all mammals with the exception of cheiroptera. Likewise no binocular vision, stereoscopic vision, stereognosis, prehensile hands, a strong pinch, all of which contributed to producing tools which in turn contributed to the control of fire, cooking, softening food, shrinking teeth, altering the oropharygeal and oral airway, contributing to speech. Perhaps among the reasons why cetaceans are sans technology.
Yes indeed LUCA is very important at a certain point in time, but it can be completely overshadowed by other things depending on what we are looking for.
If the universe was full of life, both simple and complex, but absent technologically intelligent species, it would still be a wondrous place. It would also ensure that there was no need to hide from predatory, space-faring species, and so we could explore with confidence.
But let’s examine the human features that we think are so important. Suppose the cat family developed hands with opposable thumbs? They might still have to walk on their knuckles, but would that be a problem if they can sit on their haunches and support their bodies to free up their hands for working artifacts? Is our shoulder joint so important that there is no workaround? Cats have binocular vision. Could their brains grow to enable more intelligence? Speech is an issue, but are their workarounds – like signing as we have taught apes? If signing, then writing could follow. Is it really a huge stretch to imagine intelligent, tool-using, felines as an intelligent species?
While naked, ocean-living cephalopods would have limited technology without fire, had they adapted to land living as their snail and slug cousins, they could then eventually learn to use fire. Their tentacles show sufficient dexterity to supplant hands, so I could imagine them forming some sort of intelligent species. Their use of chromatophores to communicate could be very expressive.
I wouldn’t write off alternative forms to humanoid species gaining tool-using intelligence. The universe could be full of surprising intelligences. We won’t know until we explore.
The range of motion in ungulates can come as a surprise: https://www.youtube.com/watch?v=B2eEhX70Jmw (suggest playback at 0.25x) Primate glenoid cavities are actually an average between the carnivores and ungulates.
Try to stretch out their forelimbs in the equivalent of the human coronal plane at right angles to the trunkal axis.
🏋🏻♂️🤸🏽♀️ 🧗🏿♂️
Is not the answer to life’s most persistent question “where come from I?” worth $500 billion spent over years of effort (~ 5?) when we spend $900BB _each year_ on “defense” in the USA?
“Eyes and ears” look up, humans to Mars, machines to Enceladus and other icy/ocean moons, balloons to Venus, please!
The darkest option not mentioned in the posting or comments is that if we are unique, or even if not, we are in a simulation. That begs the question of LUCA entirely:
Nick Bostrom’s argument
In his 2003 paper, Are You Living in a Computer Simulation?, Bostrom argued that the probability of us living in a simulation is close to one.
David Chalmers’ argument
In his book Reality+: Virtual Worlds and the Problems of Philosophy, Chalmers states that there is at least a 25 percent probability of us living in a simulation.
David Kipping’s research
In 2020, Kipping, an astronomer at Columbia University, found that the odds of us living in a base reality are roughly 50-50.
Digitally yours, WSG
Despite the intelligence of some of these thinkers, I tend to think that they are led astray by “logic” which rarely elucidates the truth. The tech “mind-killer” is Roko’s Basilisk.
Michael Shermer wrote “Why People Believe Weird Things” about pseudoscience. It can be extended to why intelligent people believe weird things. And I use “weird” to include any unproven statements about the world. Scientists tend to be cautious about making definitive statements because new evidence could undermine them. In my limited reading, philosophers and others who indulge in thoughts rather than experiments tend to be more dogmatic, even if their thoughts are very different from others. Hypotheses are good, but testing trumps thought as the arbiter of truth.
My bottom line on “Are we living in a simulation” is to invoke Occam’s Razor. Our universe looks like reality so accept that as the simplest explanation. If a simulation, it begs many questions. Such a “theory” requires evidence. Where is it?
The simulation argument is based on the notion of what I’d call past causality. I would say this notion is essentially religious in nature (dating back to the ‘watchmaker deity’ model), even though it is just as commonly held by atheists. Basically, it supposes that time is like a half-built wall of masonry, in which there is one firm, immutable past, one present in which some sort of bifurcation is possible, and an infinite number of possible futures, most of which do not exist. By contrast, I think that in our universe (everything we can scientifically study) there is one past, one present, and one future, all immutable; that indeterminacies in physics mean that the events during a small segment of time *could* be altered without measurably impacting the distant past or future, and that this might be the case in nearby alternate realities. I hypothesize that there are macroscopic causality violations in everyone’s routine daily life produced by the direct recall of near-future events having a ‘causal’ role in the past/current events leading up to them, meaning there is more than one possible way our particular universe ‘could have’ happened. The choice of which options actually are true would be the the basis of free will and qualia (its time inverse). These should occur in the context of a perpendicular dimension of time, beyond the reach of our physics experiments, relating the alternate universes.
At least within my idiosyncratic way of thinking about things, no finite state machine should be able to accurately simulate a human mind because it creates no paradoxes. Furthermore, even if a simulation is done using something horrible like organoid intelligence, the free will and qualia emerge within any ‘simulation’ where consciousness is experienced; the external architecture must be indeterminate and unpredictable to be able to simulate it. Wherever we observe this paradox choosing one universe over another, it is the same paradox, and the part making these meaningful free-willed decisions according to its own ‘consciousness’ is the simulated being, not the external architecture.
Hi, Alex
It appears that life arose on earth soon after it cooled off, early in its formation. What is the prevailing opinion among biologists?
a) The “early origin” suggests life will appear soon on any world with suitable conditions, which further suggests life is very common throughout the universe.
Or, alternatively…
b) The early appearance of life on this planet was a lucky accident, not likely to have been repeated elsewhere, or at least, not too often.
Clearly, the first hypothesis is much more to be preferred among astrobiologists, and particularly among SETI enthusiasts, who would prefer long periods of development conducive to higher life forms and intelligent species.
What is the gut feeling in the biological community as to the most attractive hypothesis? Is there a debate in progress, or is the field adopting a wait-and-see attitude?. Are there lines of research likely to resolve the question currently being pursued?
I can see how the discovery of extraterrestrial life would go a long way in resolving this issue, which explains the active explorations currently going on. But I have always gotten the impression that the biological community has been much more skeptical about the ubiquity of life in the cosmos than their colleagues in the physical sciences.
@Henry
I have no idea what the prevailing view amongst biologists of extraterrestrial life is. Biology is a large subject with disciplines ranging from the very small to the biosphere. Most biologists work in the areas that interest them.
But let’s remember that astrobiology is a subject that does not have any extraterrestrial biology to investigate. So it investigates peripheral subjects that may pertain to ET life.
As Drumlin says to Ellie Arroway in “Contact”, listening for ETI signals is a way to end a career. The safe route is to investigate the life and systems that exist on Earth.
As regards lines of approach. In my post in June about the ambiguity of biosignatures, Figure 4 has the 4 major lines of research that bear upon the problem. Biologists are principally involved in the 2 quadrants of research on the left, and partially in the quadrant on the upper right.
But I have to say that evidence of ET life is the most important. That is why the search for life in our system is so important. Biosignatures on exoplanets while not definitive, are important lines of inquiry. And naturally, if the SETI folks receive a signal…
Life is primarily chemistry becoming unconstrained through massive organization in multiple hierarchical levels including substantial physics in the mix.
In the presence of energy flows matter is self-organizing at the lower hierarchical levels, from which higher levels may randomly emerge.
The body-mind complex is billions of years of chemical and physical shuffling.
How many of these isolated complex lifeforms have arised and disappeared without adding to the history of life’s genome. Active regions both above and below the ocean’s surface may have developed complex life much earlier then accepted, only to be destroyed before becoming sustainable.
Isolated community of complex life may have existed 2 billion years ago.
https://newatlas.com/biology/isolated-complex-life-evolved-2-billion-years-ago/
The fossil record for ancient deep sea life’s history may be locked up in metamorphic rock someplace on the earth’s crust but most of it was destroyed in subduction. The deep surface of the ocean is still making huge amounts of crust daily from the mid-ocean ridge system, where life finds it isolated homes in geothermal activity. Oxygen produced In the Deep Sea would make a perfect place for advanced oxygen loving organisms to develop but would also disappear into the subduction zones within less then 350 million years. So the giant jigsaw puzzle of life’s genome may be missing a large number of pieces that never made it beyond their isolated strongholds.
Oxygen Produced In The Deep Sea Raises Questions About Extraterrestrial Life.
https://astrobiology.com/2024/08/oxygen-produced-in-the-deep-sea-raises-questions-about-extraterrestrial-life.html
As for how this relates to the search for exterrestrial life, all indicators point to and active surface above and below sea level for advanced life to develop and evolve.
The one type of common planet that is missing from our solar system are super earths. These types of planets would be supercharged with the active conditions that would be conducive to life.
So regarding Pre-Cambrian complex life. For the sake of argument, let us assume that the Francovillian material is evidence of complex life. Unless they emerged as a parallel line of life, they must be descendants of LUCA. What may distinguish them if they left any DNA (which they did not) is unique genes. Their fundamental biology remains the same as all other life. This is true of the many lineages that went extinct in the past that left no contemporary descendants. Their genomic information is lost.
As you know, Paul Davies’s team has been looking for what he calls “Shadow Life”. This is life that we have not recognized and which may have a very different biology as it is not a descendant of LUCA, and would be a lineage represented by the gray lines in Figure 4 except that they do have living descendants. So far that research has not found anything, and I suspect for good reason. Life is Darwinian, and the “fitter” organisms outcompete the “less fit” and eventually drive them to extinction. Sometimes these organisms can retreat to a refuge that offers them a greater fitness to survive, for example, anaerobes in an oxygen-rich planet survive where oxygen is very low or absent. We have not found and classified most species, most of which are bacteria and fungi, but occasionally a large vertebrate or plant. We may still be surprised given we have a deep ocean barely explored, and even less of the crustal biosphere.
In Advaita Vedanta (and Tibetan Buddhism) the highest stage of the Being that is subject to descriptors is the “not this” awareness. (Beyond That, it is not subject to descriptors). I sit on this chair, this chair is not I. I move these limbs, these limbs are not I. These thoughts are present, they are not I. This mind is calm (agitated, etc.), this mind is not I.
The I is awareness without an “of”. Absent any objects or events in awareness, the awareness is Consciousness, Self-awareness.
The nature of the objects (and events) presented to awarenesis, whether real or unreal, is a matter of consequence only within the realm of those objects and events: to Consciousness it is the same whether or not the rest a simulation.
Apologies for the digression… I would like to see this very useful concept updated and made more accessible (maybe!) in terms of modern neuroscience. Fear is something imposed from outside, especially via the sympathetic nervous system and fast heartrate. This is true whether it be by epinephrine, marijuana, autonomical response to the sensation produced by a central line, direct stimulation of the amygdala, or the routine functioning of the limbic system. Similarly, if you remember a place in your mind’s eye, that is the activation of specific “place cells” in the hippocampus triggering the recall of those memories. There is a beautiful episode of “Hamilton’s pharmacopoeia” where someone who mutilated themself under the influence of PCP contrasts it with other, non-NMDA/glutamate mediated drugs, saying that with the other substances he perceived a “watcher” (an unaffected part of the brain) preventing self-harm. I think it is possible, with mere introspection, to distinguish this prefrontal lobe activity in one’s own consciousness, and it does seem rather different from the rest of your thinking. So I’d say even this often-named seat of consciousness enforcing self-preservation or moral standards is part of the “not-I”. When we continue this mental dissection, I think the most central part is likely to be the thalamus, and I don’t yet have a good suggestion how to put _that_ at arm’s length. Even so, I’d say consciousness represents part of the collapse of a vast homogeneous range of superposed possibilities for all past, present and future events into a single “true” state-vector, rather than being realized directly from the physical motion, change or stimulation of the chemicals at a specific location in the neurons.
This would need testing… Let’s take the case of “Schroedinger’s mathematician”. You send him up in a highly insulated space capsule, let us say a “box”. He needs to calculate a certain number, which might be done in one of a hundred ways. Unbeknownst to him, the number has already been proved by someone else with one of those ways, and we think the others won’t work. A true (radioactive decay) random number generator unlocks just one of a hundred library files on the ship, and he starts to work using the approach favored by that one library. At a predetermined time, his ship’s computer transmits the answer back to the ground, if he has it. Ideally, the mathematician, being in a superposition of states, always sends back the right answer just like a quantum computer would – unless there is a fundamental function of his consciousness that prevents him from being in a superposed state. (But what if you ask him afterward and he says the wrong number came up? Do you need an upcoming accident with a black hole for you to get the right answer to come out now?)
In your “Schroedinger’s mathematician” thought experiment, I am not clear how they are in superimposition if they have just one way to do the calculation, preselected by chance, probably the wrong one, yet they can create the correct answer?
I can see how you are recreating the S. cat example, but is it the same? In the S. cat example, the claim is that the cat is neither/both alive or dead until the box is opened. In your thought experiment the moment that the computer sends the result, the superimposed state collapses and the result is either correct or more likely wrong (p_correct = 0.01). When asked after the box is opened, they state the same answer as was received on the ground.
Despite its famous origin, I don’t believe in superimposed states for macroscopic objects. [Although this is Douglas Adam’s joke invention of the “infinite improbability drive” in THHGTTG.] Everett’s many Worlds interpretation just posits multiverses where quantum computer computations take place. When cracking RSA encryption, the correct answer is just the one that arrives that is correct by its own self inspection. Similarly NP hard calculations return the solution as many solutions are discarded in favor of the optimum one when the superimposition of the qubits is read.
See IBM’s explanation of quantum computing What is quantum computing?
Regarding your earlier point that the future is “fixed”, doesn’t the impossibility of infinite precision mean that the future is uncertain beyond N events? If so, then there are no paradoxes to experience. But suppose there were paradoxes, without an external observer, wouldn’t one not experience a paradox simply because the change would be instantaneous, certainly at the speed of our perception? This would be similar to the experiences of the population in the movie Dark City.
Alex – I was assuming that the mathematician checks “their” own work before transmission, so that it was just a superposition of 99 different 00000… transmissions with the answer. By sending, say, a million photons circularly polarized, once per picosecond, I think it is possible to detect the 1-in-100 component of a “1” if it was sent.
The classical observation is the sticky part. I don’t see this in the IBM overview, but quantum gates normally need to be reversible in order that “garbage information”, you might say, left over from the computation won’t count as a measurement and prevent the quantum algorithm from working. Clearing all the garbage information out of the mathematician might be tricky, unless perhaps he goes in a black hole. (There’s much I don’t understand about that principle, especially the time aspect!)
The last part is purely semantic in one universe. Does the future exist “now”, or “in the future”? It will surely be certain when it happens… However, if you contrast two parallel universes, with a difference in the present, then the lack of infinite precision means that both possible present states can be compatible with the same distant past and the same distant future. This permits people to exercise free will without upending the entire plot.
The latest from John Michael Godier of the Event Horizon podcast:
The Deepening Mystery of the Last Universal Common Ancestor of Life
@Robin
What caught my attention was the basaltic glass polymerizing RNA-triphosphate into [branching?] chains. The work that I found is 2 years old so I need to follow up to see if more recent work was done.
Catalytic Synthesis of Polyribonucleic Acid on Prebiotic Rock Glasses
I wonder if an early filter is a numbers game. It could be life arises easily but unless it becomes ubiquitous across a planet it will eventually fade. On earth the battle seems to have been raging between the natural geographical trajectory of the planet and life. Life for the most part has gained the upper hand.
Mars likely did have life and now, if still there, will be in pockets of opportunity.
Unless life is able to shape geography, it will fade.
@Tesh
It you think about it, “life hanging on in refugia” is like the “Fine-Tuning” argument when applied to the Gaia Hypothesis. Because we live in a fecund world where life has occupied all the niches it can and modifies the planet to help keep the conditions stable, we tend to think that Gaia implies that life will do this…or not. But we know from evolution that complex life didn’t invade the land until the Carboniferous. Before that, it was confined to an aquatic, marine, existence. Even today, some niches are too extreme, like the Antarctic where penguins have to live within reach of the sea to find food. The glaciated tops of mountains are equally uninhabited.
When there was a “Snowball Earth”, life was similarly confined. So perhaps the idea of Gaia should be modified to: Life will expand into all possible niches that are habitable and given enough time to evolve. So consider a tidally-locked planet. Life may be confined to the terminator between the extreme heat and cold of the 2 hemispheres. Arthur C Clarke had his own variation in his Odyssey quartet, with the Europans. They evolved at subsurface ocean vents and could not migrate from them. He called this trapped between fire and ice.
So now let’s turn to Mars. Any life in existence will be in a crustal biosphere, prevented from migrating to the surface due to cold, dry, irradiated, conditions. It may spread around the planet but beneath the surface. Because the zone that is warm and wet enough is kilometers below the surface, the best way to look without extensive drilling, are impact sites where rocks have been thrown to the surface. The microbes will be dead, but accessible. Similarly, the material around the Enceladon plumes might be a good place to look for subsurface ocean life that could be deposited at the base of the plume, because surface conditions are unsuited to life.
My current thought is “Which terrestrial minerals, created by life, would still be formed on planets with different conditions to Earth?”
Hi Alex,
I guess what I was trying for (whether valid or not) is that life, simple of otherwise was likely, very quickly, ubiquitous on Earth, i.e. it spread pretty much to everywhere to all intents and purposes. From this I was inferring a couple of things (below) that would get at why is Mars (or most other places we’re started to look) devoid of obvious life.
1. (Eventual) Universal interconnective between different life forms, i.e. predator/hunted or symbiotic – pretty much across the globe
2. Point 1 being ubiquitous allows for evolution against a competitor, predator or for efficiency in hunting but also against the backdrop of the environment – again across the globe.
3. That life was pretty much everywhere made for insurance against climactic variations. The presence of different co-existing life forms (even if simple), that were interdependent, would allow a re-genesis as long as a minimum threshold was met somewhere… Having co-dependent life forms (predator/hunted or symbiotic or just being physically near enough) maximizes resource gathering. utilization and drives evolution thus closing the circle…
To your last question – I would think that Life can likely form everywhere where there is (diverse) physical substrate and an energy source. Thus Life likely formed on Mars. However, it just did not achieve the numbers to ensure gaining a balance or the upper hand over the planet’s climactic trajectory.
Firstly, competition that drives evolution is principally with members of the same species in a population. This is because they are all competing in the same niches for the same food, and in animals, for the same mates. This is the lesson of Darwin’s finches where changing climates change the food source and select finches with the appropriate beak type for the food sources. In the long term, this is what drives the radiation of species from a common ancestor. Yes, the environment has other species as predators and parasites, but generally, these are secondary.
As evolution cannot predict future changes, sufficient variation may or may not allow for a regenesis after a catastrophe such as climate change, massive volcanism, ELE impactors. The Earth does have a crustal biosphere, so theoretically, if the surface was sterilized, these microbes could eventually emerge onto teh surface. Could prokarya evolve eukarya? I have no idea. On Earth, there may not be enough time once solar output makes life impossible on the surface. Around a star with a much longer lifetime, we don’t know.
As for the big question you ask, I don’t know. That is what we hope to determine, whether in the lab, examples in the solar system, and most likely on exoplanets. The universe doesn’t care one whit what I hope or prefer, and since science works by observation and experiment, at some point it just becomes speculation. One can take extreme positions based on some “logic” or anything in between. These speculations tend to be cyclical when evidence doesn’t exist. One extreme version would be supported if we find biosignatures on exoplanets wherever we look, even planets that we might not think as suitable. Or we might find that every exoplanet in the [C]HZ that should be suitable appears sterile. All we can do is lay out possibilities and implications until we acquire hard evidence.
Coming back to Earth a moment, I was impressed by the notion that lateral gene transfer might account for the surprisingly long divergence between Bacteria and Archaea. To be sure, the “surprisingly long” wasn’t something I figured out how to evaluate statistically – what does it mean for, say, the odds that life on Earth was descended from two separate hitchhikers from one other world where life began? I have no idea. But if this reflects only the history of one pair of paralogs in a much richer population, then those odds go down.
@ MS
If both bacteria and archaea were deposited from Earth, they would probably come from the same world, otherwise their basic biology would be different, notably the genetic code. [Some bacteria do have small modifications to their genetic code compared to the standard one.]
But suppose organisms came from different worlds, with different genetic codes, but otherwise similar. Could chance swapping of genes still work in some cases if the gene sequence that was translated into a different protein had functional use in the recipient? [ Normally, frameshifts, that misread the correct codon, result in non-functional proteins that are quickly destroyed.]
Horizontal gene transfer exists in higher organisms with retroviruses that incorporate their genes into the recipients’ genome. Sometimes these are functionally useful. Humans have such retrovirus sequences that the chimps and bonobos do not. This would make our genome sequences more different than time would suggest. This is why we only use conserved, functional genes for creating phylogenetic trees, but such sequence information does have other uses.
One still unsolved mystery is how some life forms have very different life stages. For example, insects have a larval stage, then pupate, and emerge as a very different, often winged form. During the pupal stage, their bodies are almost entirely broken down and different genes are read to build them to a different body plan. Simpler cases are the transformation of motile forms to sessile ones, although this doesn’t require body breakdown, just a slow transformation. Are these organisms the result of fused genomes?
Science fiction horror has many examples of this, notably “The Quatermass Xperiment” (Hammer movie version) where some spaceborne infection during spaceflight slowly transforms one of the crew.
More controlled is the use of genes from one organism to improve the performance of another, e.g. with pest-resistant crops. Because of the plant reproduction via pollen, these modified plants can “infect” the same species that are nearby. How many natural cross-fertilizations happen in close-enough plant species, or even aquatic animals that use external fertilization?
All known organisms have much in common – for example, the ribosome is a tremendously complex machine, made from two pieces of rRNA, decorated by a large number of highly conserved proteins, working with similar partners (tRNA), doing the same job of protein synthesis. I don’t think it’s plausible to have evolved it on two different worlds.
At an earlier stage, abiogenesis might have more compatibility, and it might allow for some gene transfer of catalytic RNA. If two worlds both have formose reactions, and polyphosphates, maybe they both got to a stage of nucleotides. I think Earth had more diversity at the start than we normally imagine on nucleotides, with things like riboflavin and nicotinamide perhaps being counted among the bases; any planet developing this mechanism could have started with similar diversity. Question is… is it inevitable that our life was restricted to A, C, G, T because they could form a consistent system of self-replication, or more of an accident? It seems at least conceivable that evolution might replay to compatible RNA sequence on many worlds. However, the meaning of that when translated into protein, though highly conserved on Earth, is still arbitrary — early experiments were able to change which amino acid would be made by a particular tRNA. We keep these meanings for the same reason we don’t change the meaning of the letter “r” in English – too much is written already. (The closest approach to doing this are fMet and amber/ochre/umber mutant bacteria, but it’s not that impressive really)
If the RNA were inevitable, and maybe the biochemistry is inevitable, then aliens might be close enough to earthlings to be able to eat them, if they’ve evolved robust resistance to toxins. The smart money would bet against it, but it does make for a more energetic story. It is even somewhat conceivable that an alien microorganism could have landed on Earth, become associated with one of our lifeforms as an endosymbiote, and still be hiding out in the cells of that organism today, despite (presumably) failing to survive the evolutionary contest on its own. I wouldn’t bet on it, but the fun part of biology is you never really know.
Ecdysis isn’t really that special. Humans shed skin ranging from single cells to little flecks of dandruff, but snakes, with harder scales, make a go of it in a single moult. Ecdysozoans (butterflies, for example) are a more extreme example. Human skin cells slowly move up and form a granular layer as permitted under the influence of thyroid hormone, which hardens somewhat. Butterflies have ecdysone, a smarter replacement for thyroid hormone, and they use it differently, to cause all the undifferentiated skin to grow at once as the differentiated skin is prepared to be shed. The wings of a butterfly are hidden away inside the caterpillar as “imaginal disks”, in little recesses of the skin, and so they make a striking appearance when they finally can grow large, becoming visible on the outside of a pupa. Even more so when they grow gigantic and dominate the appearance of the adult. But it’s all part of a process in which the main thing that changes is the relative sizes of the structures. Similarly, the difference between male and female humans is much less than people imagine – just a few key switches thrown to make the overall form come out differently. It isn’t really a chromosome’s worth of difference; that evolves only as a matter of genetic gamesmanship, with one gene (SRY) able to do the largest part of the function of the whole Y chromosome. The transformation of a tunicate is a bit more striking though – humans and other vertebrates are fortunate to have lost their adult stage.
Jeremy England’s book Every Life on Fire: How Thermodynamics Explains the Origins of Living Things attempts to bridge the gap between physics & chemistry on one hand and biology on the other.
“Every Life Is on Fire: How Thermodynamics Explains the Origins of Living Things” by England, Jeremy
Clay has been suggested as a substrate and a template for the assembly and polymerization of nucleotides in the formation of early RNA.
Incidentally Adam refers to origin in the ground
Alternative Pathways in Astrobiology: Reviewing and Synthesizing Contingency and Non-Biomolecular Origins of Terrestrial and Extraterrestrial Life
https://astrobiology.com/2024/09/alternative-pathways-in-astrobiology-reviewing-and-synthesizing-contingency-and-non-biomolecular-origins-of-terrestrial-and-extraterrestrial-life.html
The paper effectively takes teh diagram in my Figure 4 and extrapolated it with speculation about pre-biotic chemistry. While their speculations may be true, there is no evidence for it, nor the “contingency” hypothesis. [Nor can there be unless a demonstrated in the lab. I am not even sure that it could be tested or falsified using prebiotic exoplanet samples, entirely different from finding life with a different biology to earth life.]
https://astrobiology.com/2024/09/astrobiology-mission-update-juice-confirms-that-earth-is-habitable.html
Astrobiology Mission Update: Juice Confirms That Earth Is Habitable
By Keith Cowing
Press Release
ESA
September 10, 2024
During its flyby of Earth on 20 August, ESA’s Jupiter Icy Moons Explorer (Juice) found ingredients for life in Earth’s atmosphere.
The flyby provided the opportunity to test out and calibrate Juice’s science instruments in space, ensuring that they are ready for arrival at Jupiter.
Two of Juice’s instruments – the Moons and Jupiter Imaging Spectrometer (MAJIS) and the Submillimetre Wave Instrument (SWI) – collected data that confirm Earth is habitable.
In this context, we’re defining ‘habitability’ as having conditions necessary to life to arrive and survive. Jupiter’s icy moons are particularly exciting prospects for life considering the oceans that hide beneath their surfaces.
During the Earth flyby, SWI ‘listened’ for the signals from hundreds of molecules in Earth’s atmosphere, including water and the so-called ‘CHNOPS’ elements (carbon, hydrogen, nitrogen, oxygen, phosphorous and sulphur). The CHNOPS elements are the most common ingredients of living organisms.
Atmospheric composition measured by ESA’s Jupiter Icy Moons Explorer’s (Juice) Submillimetre Wave Instrument (SWI) on 20 August, during the mission’s Earth flyby. The instrument was looking straight down through the atmosphere towards Earth’s surface (indicated by the blue circle that tracks over Earth’s surface). — ESA
MAJIS also measured the composition of the atmosphere, detecting important molecules such as oxygen, ozone, carbon dioxide and water. Furthermore, the instrument imaged Earth’s surface in infrared light, resulting in information-rich temperature maps.
The MAJIS team is preparing to dig further into the data they collected during the Earth flyby, especially on the concentration of oxygen in the atmosphere. Their findings will help them discover whether the levels of oxygen would support current biological activity on Earth.
MAJIS detects different molecules in Earth’s atmosphere — ESA
ESA Juice project scientist Olivier Witasse says: “We are obviously not surprised by these results… it would have been extremely concerning to find out that Earth was not habitable! But they indicate that MAJIS and SWI will work very successfully at Jupiter, where they will help us investigate whether the icy moons could be potential habitats for past or present life.”
MAJIS thermal infrared image of the Indian Ocean and Malaysia — ESA
Once at Jupiter, SWI will study the composition of the planet and its icy moons, telling us more about their current climates, their origin, and their history. The information that the instrument collects will reveal not only their potential habitability, but also capture signs of their current biological activities.
Meanwhile, MAJIS will observe Jupiter’s clouds and the ingredients in its atmosphere, and will investigate the thin atmospheres and ices and minerals on the surfaces of the icy moons.
SWI was developed and built by an international consortium of institutes led by the Max Planck Institute for Solar System Research (Germany). This development was supported by national funding agencies including DLR (Germany).
MAJIS was developed by a large consortium of European scientists and engineers, with two major contributions by France and Italy through the support of their respective space agencies, CNES and ASI. The main scientific contribution and responsibility are from the Institut d’Astrophysique Spatiale (France) and the National Institute for Astrophysics (Italy).
https://astrobiology.com/2024/09/oceans-and-continents-devastated-marine-life-millions-of-years-ago.html
Oceans And Continents Devastated Marine Life Millions Of Years Ago
By Keith Cowing
Press Release
University of Southampton
September 11, 2024
Scientists have revealed how a “tag-team” between the oceans and continents millions of years ago devastated marine life – and altered the course of evolution on Earth.
Their study has unearthed a new explanation for a string of severe environmental crises, called oceanic anoxic events, which happened between 185 and 85 million years ago.
These occurred when the seas became critically depleted of dissolved oxygen.
Experts from the University of Southampton, which led the study, said these events triggered significant biological upheavals, including mass extinctions of marine species.
The findings are published today Thursday 29 August in Nature Geoscience.
Lead author Tom Gernon, a Professor of Earth Science at Southampton, said: “Oceanic anoxic events were like hitting the reset button on the planet’s ecosystems.
“The challenge was understanding which geological forces hit the button.”
The study was undertaken by Southampton in collaboration with academics from the universities of Leeds, Bristol in the UK, Adelaide in Australia, Utrecht in the Netherlands, Waterloo in Canada, and Yale in the US.
The researchers examined the impact of plate tectonic forces on ocean chemistry during the Jurassic and Cretaceous Periods, collectively known as the Mesozoic era.
This chapter of Earth’s history is often dubbed the age of the dinosaurs, said Prof Gernon, and is famously exposed along the Jurassic Coast on the UK’s south coast as well as along the cliffs of Whitby in Yorkshire and Eastbourne in East Sussex.
The team combined statistical analyses and sophisticated computer models to explore how chemical cycles in the ocean could have feasibly responded to the breakup of the supercontinent Gondwana, the great landmass once roamed by the dinosaurs.
Prof Gernon added: “The Mesozoic era witnessed the breakup of this landmass, in turn bringing intense volcanic activity worldwide.
“As tectonic plates shifted and new seafloors formed, large amounts of phosphorus, a nutrient essential for life, were released from weathering volcanic rocks into the oceans.
“Crucially, we found evidence of multiple pulses of chemical weathering on both the seafloor and continents, which alternately disrupted the oceans.
a,b, Plate-tectonic reconstructions (Methods) showing the main palaeogeographic features, MORs and exposed large igneous provinces, as well as the approximate distribution of OAE-related sequences in the Toarcian OAE at about 183–182 Ma (with OAE sites from ref. 49 and references therein) (a) and Turonian at about 90 Ma (with OAE sites from ref. 50 and references therein) (b). Note that shallow seas include epicontinental seaways, including the Western Interior Seaway of North America. CIE, carbon isotope excursion; HALIP, High Arctic Large Igneous Province. Figure created with GPlates26 (https://www.gplates.org/). — Nature Geoscience
“It’s like a geological tag-team,” said Prof Gernon.
Experts from the universities found the timing of these weathering pulses matched up with most oceanic anoxic events in the rock record.
They propose that the weathering-related influx of phosphorus to the ocean acted like a natural fertiliser, boosting the growth of marine organisms.
However, the researchers said these fertilisation episodes came at a major cost for marine ecosystems.
The increase in biological activity led to huge amounts of organic matter sinking to the ocean floor, where it consumed large quantities of oxygen, said co-author Benjamin Mills, a Professor of Earth System Evolution at the University of Leeds.
He added: “This process eventually caused swathes of the oceans to become anoxic, or oxygen-depleted, creating ‘dead zones’ where most marine life perished.
“The anoxic events typically lasted around one to two million years and had profound impacts on marine ecosystems, the legacy of which are even felt today.
“The rocks rich in organic matter that accumulated during these events are by far the largest source of commercial oil and gas reserves globally.
As well as explaining the cause of extreme biological turmoil in the Mesozoic, the study’s findings highlight the devastating effects that nutrient overloading can have on marine environments today.
The team of researchers xplained how present-day human activities have reduced mean oceanic oxygen levels by about two per cent – leading to a significant expanse in anoxic water masses.
Prof Gernon added: “Studying geological events offers valuable insights that can help us grasp how the Earth may respond to future climatic and environmental stresses.”
Overall, the team’s findings reveal a stronger-than-expected connection between the Earth’s solid interior and its surface environment and biosphere, especially during periods of tectonic and climatic upheaval.
“It’s remarkable how a chain of events within the Earth can impact the surface, often with devastating effects”, added Prof Gernon.
“Tearing continents apart can have profound repercussions for the course of evolution”.
Solid Earth forcing of Mesozoic oceanic anoxic events, Nature Geoscience (open access)
https://www.cnn.com/2024/09/20/science/human-genome-crystal-intl-scli/index.html
Scientists store entire human genome on ‘memory crystal’ that could survive billions of years
By Rosa Rahimi, CNN
Published 7:53 AM EDT, Friday, September 20, 2024
Scientists in the United Kingdom have stored the entire human genome on a “5D memory crystal,” in the hope that it could be used in the future as a blueprint to bring humanity back from extinction.
The crystal, which was developed by a team of researchers at the University of Southampton’s Optoelectronics Research Centre, could also be used to create a record of plant and animal species faced with extinction.
It can hold up to 360 terabytes of information for billions of years and can withstand extreme conditions, including freezing, fires, direct impact force, cosmic radiation and temperatures of up to 1,000 degrees Celsius, the university said in a press statement published Thursday.
In 2014, the crystal was awarded the Guinness World Record for “most durable digital storage material.”
Kazansky’s team used ultra-fast lasers to inscribe the human genome data into voids as small as 20 nanometers (a nanometer is about one-billionth of a meter).
They describe the data storage on the crystal as 5D because the information is translated into five different dimensions of its nanostructures — their height, length, width, orientation and position.
“The 5D memory crystal opens up possibilities for other researchers to build an everlasting repository of genomic information from which complex organisms like plants and animals might be restored should science in the future allow,” said Peter Kazansky, a professor of optoelectronics, who led the team at Southampton.
The team had to consider who – or what – would retrieve the information, so far off into the future.
It could be an intelligence (species or machine) – or it could be found in a future so distant that no frame of reference would exist for it. To help whoever finds it, the researchers included a visual key.
“The visual key inscribed on the crystal gives the finder knowledge of what data is stored inside and how it could be used,” said Kazansky.
“Their work is super impressive,” said Thomas Heinis, who leads research on DNA storage at Imperial College London and was not involved in the study. However, he says questions remain about how such data could be read in the future.
“What Southampton presents probably has a higher durability, however, this begs the question: what for? Future generations? Sure, but how will they know how to read the crystal? How will they know how to build the device to read the crystal? Will the device be available in hundreds of years?” he added. “I can barely connect my 10-year-old iPod and listen to what I listened back then.”
For now, the crystal is stored in the Memory of Mankind archive, a time capsule within a salt cave in Austria.
In 2018, Kazansky and his team used the memory crystal technology to store Isaac Asimov’s “Foundation” trilogy of science fiction books, which were then launched into space aboard a Tesla Roadster. The technology has also been used to store major documents from human history, including the Universal Declaration of Human Rights and the Magna Carta.
https://www.southampton.ac.uk/news/2016/02/5d-data-storage-update.page
Earlier this year, scientists revealed a plan to safeguard Earth’s species in a cryogenic biorepository on the moon, intended to save species in the event of a disaster on our home planet.
https://www.cnn.com/2024/08/05/science/cryogenic-repository-moon-scli-intl/index.html